Меню
Меню





 Все темы
Все темы
Ферменты плазмодия, расщепляющие лизолецитины хозяина, могут стать мишенью противомалярийных препаратов
Поражая эритроциты хозяина, малярийный плазмодий получает необходимые ему жирные кислоты за счет гидролиза лизофосфатидилхолинов (лизолецитинов) сыворотки крови. Ученые определили два фермента, с помощью которых плазмодий это делает, — XL2 и XLH4. Без них плазмодий не получает жирных кислот и страдает от токсичности лизофосфатидилхолинов. Ингибирование этих ферментов может стать новым подходом к борьбе с малярийным плазмодием.
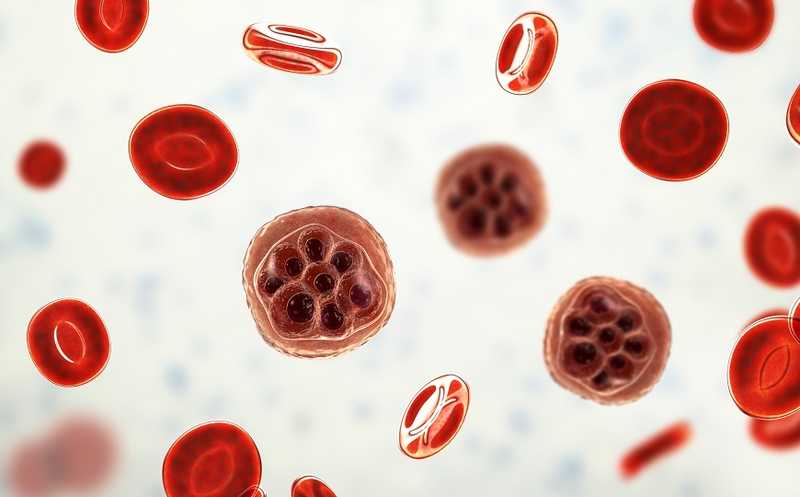
Во время бесполой стадии своего жизненного цикла малярийный плазмодий заражает эритроциты человека. В этот период плазмодию необходимо много фосфолипидов, из-за резкого увеличения площади мембран паразита. Хотя плазмодий и способен сам синтезировать жирные кислоты, из которых можно получить фосфолипиды, он также восполняет свои нужды за счет хозяина. В качестве источника жирных кислот плазмодий использует свободные жирные кислоты и лизофосфатидилхолины (ЛФХ, лизолецитины), содержащиеся в сыворотке крови. ЛФХ могут поддерживать рост паразита, даже являясь единственным источником жирных кислот. Американские ученые решили выяснить, как именно малярийный плазмодий метаболизирует ЛФХ.
Сначала исследователи определили, использует ли плазмодий собственные ферменты или ферменты эритроцитов хозяина для гидролиза ЛФХ. Они разработали тест in situ, позволяющий оценить гидролиз ЛФХ в зараженных эритроцитах. Ученые внесли флуоресцентную метку в церамиды (жирные кислоты, связанные со сфингозином), которые в плазмодии преобразовывались в меченые жирные кислоты. Впоследствии паразит синтезировал из них нейтральные липиды. Авторы исследования предположили, что флуоресценция от полученных таким образом липидов будет гаситься по мере образования жирных кислот из ЛФХ, конкурирующих с мечеными жирными кислотами, и это предположение оказалось верным.
Затем ученые проводили тот же тест с использованием различных малых молекул — ингибиторов сериновых гидролаз. Наиболее эффективным ингибитором оказалось вещество под названием AKU-010. При этом ингибирующий эффект наблюдался в зараженных эритроцитах, но не в здоровых. Так ученые сделали вывод о том, что паразит сам вырабатывает ферменты, которые гидролизуют ЛФХ.
Используя протеомные данные, исследователи идентифицировали несколько кандидатных ферментов, выполняющих гидролиз ЛФХ. Первые два из них назвали экспортируемыми липазами — XL1 и XL2, так как они выходили из клетки плазмодия в эритроцит. С помощью CRISPR/Cas9 ученые сделали три линии с нокаутами генов гидролаз: ΔXL1, ΔXL2, ΔXL1/2. Это позволило выяснить, что XL2 экспрессируется плазмодием в бо́льших количествах, чем XL1. Однако нокаут обоих этих ферментов снизил уровень гидролиза ЛФХ лишь незначительно, то есть плазмодий может осуществлять его и другими способами.
Дальнейшие поиски обнаружили еще две сериновые гидролазы, которые исследователи назвали «гомологами экспортируемых липаз» — XLH3 и XLH4. В отличие от XL1 и XL2, они находятся внутри паразита и не выходят в эритроциты. При этом XLH3 практически не экспрессируется на бесполой стадии жизни плазмодия.
С помощью CRISPR/Cas9 ученые сгенерировали еще несколько нокаутных линий плазмодия, чтобы проверить роль этих липаз, — ΔXLH4, ΔXL2/XLH4, ΔXL1/2/XLH4, а также ΔXL1/2/XLH3/4. Потеря только XLH4 оказала минимальное влияние на уровень гидролиза ЛФХ, а в линии ΔXL2/XLH4 он значительно снизился. Следовательно, для гидролиза ЛФХ плазмодий использует главным образом ферменты XL2, который экспортируется в эритроцит, и XLH4, который остается внутри плазмодия.
Ученые подтвердили эти результаты, проведя тест на лизофосфолипазную активность ферментов с флуоресцентно меченными ЛПХ. При потере только XL2 лизофосфолипазная активность лизатов зараженных эритроцитов падала в шесть раз, при потере XLH4 — в три раза. При этом комплементация XL2 в линиях, в которых этот белок отсутствовал, восстанавливала активность.
Далее исследователи выращивали нокаутных плазмодиев в среде, в которой ЛПХ были единственным источником жирных кислот. Они не могли выживать в этой среде, хотя в предыдущих исследованиях было показано, что пальмитата и олеата достаточно для поддержания роста на протяжении нескольких поколений. При комплементации XL2 их рост восстанавливался.
Ученые предположили, что лизофосфолипазы необходимы паразитам как для выработки жирных кислот, из которых синтезируются липиды, так и для защиты от токсического действия ЛПХ. Действительно, нокаутные плазмодии не могли расти и размножаться ни в среде, содержащей только ЛПХ, ни в среде с 10% сыворотки крови, которая содержит и свободные жирные кислоты, и ЛПХ. Тем не менее рост плазмодиев всегда возобновлялся при комплементации XL2.
При действии на зараженные эритроциты AKU-010, ингибирующего XL и XLH, наблюдался тот же эффект, что и при нокауте генов соответствующих ферментов. Следовательно, ингибирование ферментов, с помощью которых плазмодий проводит гидролиз ЛПХ, может стать новой стратегией для борьбы с малярией. Однако AKU-010 и похожие ингибиторы могут оказаться чрезмерно токсичными для медицинского применения, поэтому ученым предстоит модифицировать эту малую молекулу для снижения токсичности.
Длинная некодирующая РНК малярийного плазмодия необходима для его размножения
Источник
Liu, J. Metabolism of host lysophosphatidylcholine in Plasmodium falciparum-infected erythrocytes. // PNAS, 121(8) (2024). DOI: 10.1073/pnas.2320262121
Препринт на biorXiv





 0
0