Меню
Меню





 Все темы
Все темы
Анфолдаза p97 обеспечивает разрушение патогенных белковых агрегатов
При болезнях Альцгеймера, Паркинсона и других нейродегенеративных заболеваниях в клетках мозга накапливаются белковые агрегаты. Авторы статьи в Nature Communications исследовали процесс разрушения агресом — внутриклеточных контейнеров для агрегатов. Ключевую роль в нем играет убиквитинзависимая анфолдаза p97, обеспечивающая фрагментацию и выведение агресом. Понимание этого процесса открывает новые пути для разработки лекарств, способных предотвратить или замедлить нейродегенерацию.
Nat Commun 2025. DOI: 10.1038/s41467-025-59556-x | CC BY 4.0
В клетках животных существует множество систем контроля качества белков (PQC), которые регулируют их обмен. Но при клеточных стрессах работа этих систем может нарушаться, и начинают формироваться белковые агрегаты. Агрегирование белков приводит к развитию таких патологий, как болезнь Альцгеймера, болезнь Паркинсона, хорея Гентингтона, и некоторых видов рака. Чтобы временно нейтрализовать белковые агрегаты, клетка помещает их в агресомы — включения, ограниченные снаружи белком виментином. Затем они подвергаются селективной аутофагии, и обмен белка восстанавливается. В то время как процесс формирования агресом изучен, детали их дезинтеграции остаются неясны. Ученые из Вюрцбургского университета в Германии с коллаборатором из Гарварда выявили ключевых участников этого процесса — это убиквитинселективная анфолдаза p97 (альтернативное название VCP) и белок-шаперон Hsp70.
При помощи флуоресцентной микроскопии исследователи выявили надежный маркер агресом — рецептор селективной аутофагии TAX1BP1. Его использовали для анализа белков, входящих в агресому. Для этого к нему генетически «пришивали» биотинлигазу, и она помечала биотином все белки, находящиеся поблизости TAX1BP1. Среди них оказались протеосома 26S, p97, шапероны и ряд других белков (AMOTL2, PLEK2, FAM83D и PFN2).
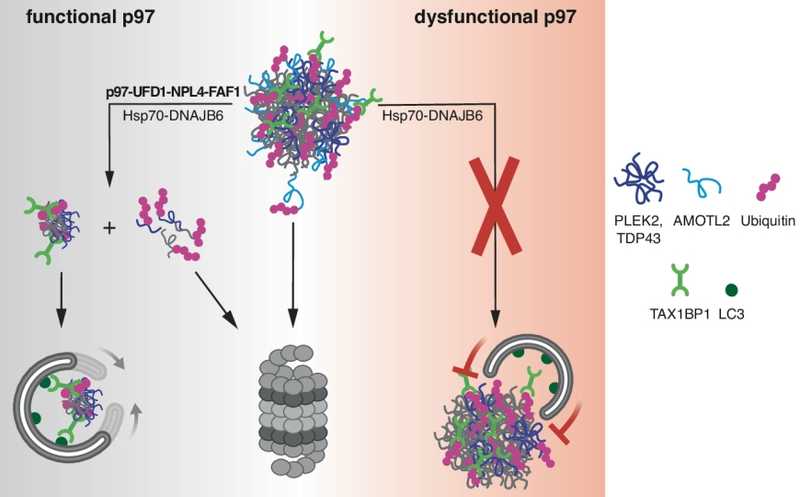
Серия экспериментов с ингибированием p97 и Hsp70 показала их роль в дезинтеграции агресом. Сперва в клетках было химически индуцировано накопление агрегатов, затем в течение 15 часов клетки восстанавливались, и после этого их инкубировали с ингибиторами PQC в течение 6 часов. В клетках, обработанных ингибитором p97, выведение агресом было нарушено сильнее всего. Обработка ингибитором Hsp70 также значительно снижало их дезинтеграцию, в отличие от ингибиторов лизосомальной деградации и протеосом, которые оказывали менее заметные эффекты.
С помощью флуоресцентной микроскопии авторы идентифицировали ряд кофакторов p97, локализованных в агресомах: UFD1, NPL4, UBXN1, FAF1, PLAA и HOIP. А среди белков из окружения агресомного маркера TAX1BP1 дополнительно выявили кофактор VCPIP1. Ингибирование комплекса UFD1-NPL4 методом РНК-интерференции существенно нарушало утилизацию агресом, но не влияло на их формирование. Блокирование домена цинковых пальцев в комплексе оказывало тот же эффект. Снижение уровня белков FAF1 и PLAA в клетках также тормозило клиренс агресом, при этом снижение FAF1 (как и UFD1-NPL4) ухудшало агресомную локализацию p97. Недостаток других кофакторов на утилизацию агресом не влиял. Таким образом, UFD1-NPL4 и FAF1 необходимы для рекрутирования p97 к агресомам и расщепления агрегированных белков в ней.
Авторы статьи изучили механизм расщепления белков, накапливающиеся в агресомах, таких как AMOTL2 и PLEK2. Было установлено, что AMOTL2 преимущественно деградирует протеасомами независимо от p97 и Hsp70, в то время как расщепление нерастворимого PLEK2 требует участия протеасом, p97, Hsp70 и аутофагии. Белок FAM83D, подобно PLEK2, частично утилизируется через аутофагию, вероятно, в качестве «попутчика» агрегированных белков. Кроме того, исследователи установили, что агрегированная мутантная форма TDP-43, связанная с нейродегенеративными заболеваниями, является p97-зависимым белком агресом и частично деградирует через аутофагию.
Чтобы выяснить, почему p97 необходим для клиренса агресом, авторы статьи исследовали, как формируются агрефагосомы – аутофагосомы, содержащие агресомы, — положительные по маркерам аутофагии (убиквитину и LC3). Ингибирование p97 приводило к нарушению фрагментации агресом и, как следствие, к снижению количества агрефагосом, но при этом маркеры селективной аутофагии p62 и TAX1BP1 нормально локализовались на агресомах, а формирование аутофагосом не были нарушены. Методом микроскопии сверхвысокого разрешения было показано, что в норме LC3 ассоциируется с фрагментами распадающихся агресом, формируя агрефагосомы, в то время как при ингибировании p97 LC3 практически не обнаруживался на стабильных агресомах. Следовательно, p97-опосредованная дезинтеграция агресом необходима для их последующей селективной аутофагии.
Эти результаты углубляют понимание механизмов клеточного ответа на стресс и могут способствовать разработке новых терапевтических подходов для борьбы с нейродегенеративными и другими заболеваниями, связанными с накоплением белковых агрегатов. Исследование подтверждает, что мутации, нарушающие функции p97/VCP, нарушают деградацию агресом и могут вносить вклад в развитие некоторых форм деменции и нейромышечных дегенеративных заболеваний, отмечается в пресс-релизе.
RING-Bait разбирает агрегаты тау-белка и улучшает двигательную функцию у мышей с таупатией
Источник
Maria Körner, et al. p97/VCP is required for piecemeal autophagy of aggresomes // Nature Communications 16, 4243. Published online 7 May 2025. DOI: 10.1038/s41467-025-59556-x
Вам будет интересно
 113
113
 0
0