Меню
Меню





 Все темы
Все темы
Источники патогенного ApoE4 в мозге — пограничные макрофаги
Нейродегенеративное действие ApoE4, патогенной изоформы аполипопротеина E, обусловлено микрососудистыми повреждениями ткани мозга. ApoE вырабатывают различные типы клеток в мозге, однако новое исследование показало, что нейроваскулярную дисфункцию вызывает только ApoE4, который производят пограничные макрофаги мозга. Интересно, что они же являются его эффекторами — в ответ на появление ApoE4 продуцируют активные формы кислорода.
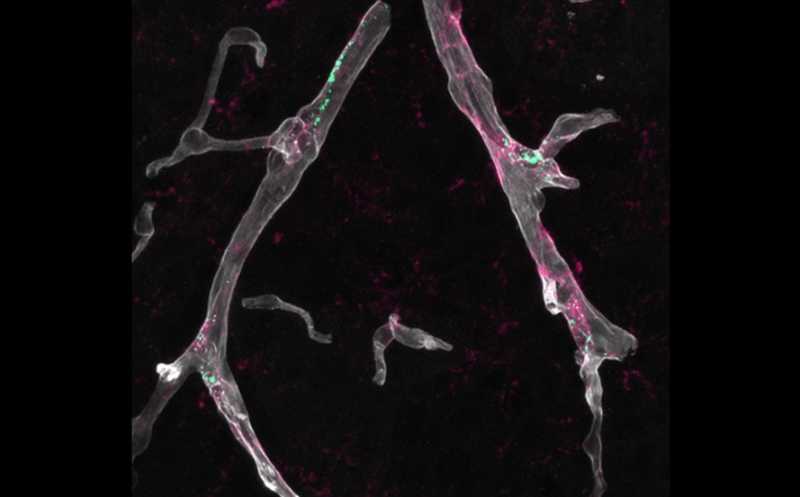
Пограничные макрофаги мозга (зеленые) производят белок ApoE (пурпурный) — в данном случае патогенную изоформу ApoE4, ассоциированная с повышенным риском нейродегенерации. Это способствует повреждению кровеносных сосудов (серые).
Credit:
Park and Iadecola labs | Пресс-релиз
Аполипопротеин Е, или ApoE — белок, опосредующий обмен липидов (триглицеридов и холестерина). Ген ApoE характеризуется наличием полиморфизмов, которые формируют основные аллельные варианты и соответствующие изоформы белка. Один из вариантов — ApoE4 — ассоциирован с повышенным риском нейродегенерации, ведущей к болезни Альцгеймера. Существуют данные, согласно которым ApoE4 повышает риск микрососудистых патологий и нарушает кровоснабжение мозга. Однако до сих пор неясно, какой тип клеток продуцирует ApoE4 в головном мозге, и механизмы его повреждающего воздействия точно не известны.
Основной причиной деменции считают микрососудистое повреждение подкоркового белого вещества, которое особенно уязвимо для подобных нарушений. Один из компонентов нейроваскулярной сети— пограничные макрофаги (border-associated macrophages, BAM): особый тип миелоидных клеток головного мозга, которые присутствуют в мозговых оболочках, сосудистом сплетении и периваскулярных пространствах. Они, в частности, способствуют очищению лимфатической системы мозга, ограничивают распространение инфекций. Роль пограничных макрофагов в развитии нейроваскулярной дисфункции была показана на животных моделях болезни Паркинсона, инсульта, гипертензии. Авторы новой статьи в Nature Neuroscience поставили перед собой задачу изучить механизм пагубного действия ApoE4 и найти источник этого белка в головном мозге. Выяснилось, что этот белок тесно связан с пограничными макрофагами: они и производят его, и являются его мишенью.
Чтобы выяснить, как ApoE4 влияет на кровообращение мозга, мышам наносили раствор этого белка на неокортекс местно, через отверстие в черепе. Изменения мозгового кровотока регистрировали с помощью лазерной допплеровской флоуметрии; например, в участке коры, связанным с вибриссами, кровоток увеличивался в ответ на прикосновение к вибриссам. Местное введение рекомбинантного человеческого ApoE4 (rApoE4) мышам дикого типа уменьшило индуцированное повышение кровотока. А в аналогичном эксперименте с рекомбинатным человеческим ApoE3 (rApoE3; этот вариант считается доброкачественным, не влияющим на риск нейродегенеративных заболеваний) такого эффекта не было.
Нейроваскулярную дисфункцию, вызванную ApoE4, ослаблял ингибитор NOX — NADPH-оксидазы, продуцирующей активные формы кислорода (АФК). Таким образом, ApoE4 способствует образованию АФК и инициации окислительного стресса.
С целью поиска клеточного источника АФК мышам дикого типа вводили флуоресцирующий декстран (Alexa Fluor-680–декстран), который поглощают пограничные макрофаги, и маркер АФК дигидроэтидин. На таких мышах повторяли эксперименты с нанесением на неокортекс rApoE4 и rApoE3. Визуализация с помощью двухфотонного микроскопа продемонстрировала повышение АФК-зависимой флуоресценции в пограничных макрофагах неокортекса в присутствии rApoE4, но не rApoE3. Следовательно, только изоформа ApoE4 запускает выработку АФК пограничными макрофагами.
Затем в неокортекс мышам дикого типа ввели клодронат, избирательно разрушающий макрофаги, и после этого у них ApoE4 не снижал мозговой кровоток, что еще раз подтвердило его действие именно через этот тип клеток.
Из литературы известно, что пограничные макрофаги могут быть и источником ApoE4. Чтобы проверить эту возможность, авторы, используя Cre-рекомбиназу, индуцируемую тамоксифеном, создали мышиную модель Mrc1Cre+/ ApoE4fl/fl. В организме таких мышей экспрессируется патогенный ApoE4, но если им ввести тамоксифен, у них избирательно выключается экспрессия гена ApoE4 в пограничных макрофагах. Действительно, тамоксифен нормализовал у мышей нейрососудистые реакции. Однако при введении rApoE4 в их неокортекс ученые снова наблюдали нейрососудистую дисфункцию. Это означает, что пограничные макрофаги — по-видимому, основные источники ApoE4.
Затем авторы показали, что пограничные макрофаги являются и эффекторами дисфункции, вызванной ApoE4. Мышам, у которых их собственный ген ApoE был заменен одним из вариантов человеческого – ApoE3 или ApoE4, облучали головной мозг, чтобы истощить пул пограничных макрофагов, а затем трансплантировали костный мозг мыши другой линии. В результате получили химер — мышей с геном ApoE3 и пограничными макрофагами, экспрессирующими ApoE4, а также мышей с геном ApoE4 и ApoE3-продуцирующими макрофагами. ApoE4-макрофаги индуцировали нейроваскулярную дисфункцию даже у мышей с геном доброкачественного ApoE3, тогда как ApoE3 -акрофаги у ApoE4-мышей ее устраняли.
Это исследование делает более полным наше понимание патогенного действия ApoE4. Как оказалось, этот белок индуцирует продукцию АФК в пограничных макрофагах, более того, основным источником ApoE4 являются сами пограничные макрофаги. Полученные данные имеют огромное значение для понимания нейроваскулярной патологии болезни Альцгеймера — неизлечимого на данный момент нейродегенеративного заболевания.
Источник
Anfray, A. et al. A cell-autonomous role for border-associated macrophages in ApoE4 neurovascular dysfunction and susceptibility to white matter injury. Nature Neuroscience (2024), published online 18 september 2024. DOI: 10.1038/s41593-024-01757-6

Вам будет интересно



 160
160
 0
0