Меню
Меню





 Все темы
Все темы
Асимметрию деления растительных клеток обеспечивает пространственное ограничение цитоскелета
Асимметричное деление необходимо для образования специализированных клеток, в том числе клеток устьиц у растений. Новая статья в Science описывает механизмы асимметричного деления у Arabidopsis thaliana. Ключевое значение при этом имеют полярные комплексы BASL/BRXf, которые «запрещают» деление, проходящее через определенный участок мембраны.
Для дифференциации клеток самых разных многоклеточных организмов необходимо их асимметричное деление, при котором генетические детерминанты, определяющие дальнейшую клеточную судьбу, попадают лишь в одну из дочерних клеток. У животных асимметричность взаимодействием определенных детерминант с цитоскелетом, однако у растений механизм асимметричности обнаружить не удалось. Такие механизмы впервые описали авторы новой статьи в Science на примере эпидермальных клеток-предшественниц устьиц Arabidopsis thaliana. Они дают начало замыкающим клеткам, которые управляют газообменом растения, открывая и закрывая устьичную щель за счет изменения тургора.
Известно, что асимметричному клеточному делению у растений предшествует формирование так называемых полярных комплексов, связанных с клеточной мембраной. (Полярность в широком смысле — асимметричное распределение тех или иных компонентов внутри клетки.) За это отвечают белки BASL (breaking of asymmetry in these stomatal lineage) и белковое семейство brevis radix (BRXf). Перед митозом ранний комплекс BASL/BRXf рекрутирует другой белок с полярной локализацией — POLAR, а тот в свою очередь привлекает GSK3-подобные киназы, стимулирующие асимметричное деление. Комплекс BASL/BRXf занимает определенный участок мембраны, называемый «полумесяцем полярности» (polarity crescent).
После того, как предшественницы замыкающих клеток получают комплекс BASL/BRXf от родительских клеток, он активирует регуляторный путь MAPK-киназ и тем самым подавляет способность клеток оставаться в состоянии меристемы (неограниченного деления).
Авторы проследили развитие семядолей A. thaliana с флуоресцентно меченными ядрами, плазматическими мембранами и полярными комплексами BASL/BRXf. Удалось подтвердить, что асимметричное деление всегда связано с попаданием BRXL2 в одну из двух дочерних клеток.
Более детальное рассмотрение обнаружило два разных класса клеток с асимметричным делением. В 73% случаев митоз происходил по линии, соответствующей кратчайшему расстоянию между противоположными клеточными стенками, на которой также находилось ядро. Такое деление характерно для большинства изученных растительных клеток. В этом случае асимметричным делением управляет смещение ядра, регулируемое полярным комплексом, и минимизация площади плоскости деления.
Второй класс асимметричного деления отмечен у 27% клеток: оно идет в плоскости, резко отличной от минимальной по площади. Это предполагает участие других механизмов регуляции. Подобное клеточное деление наблюдается не только у эпидермальных клеток-предшественниц устьиц, однако в других тканях A. thaliana оно связано с внешней регуляцией — например, механическими взаимодействиями тканей или фитогормонами.
Авторы решили узнать, насколько необходима полярность, зависимая от BASL/BRXf, для появления клеток с разными классами асимметричного деления. Оказалось, что при потере полярности из-за мутаций в basl эти классы становятся неразличимы, следовательно, поляризация BASL/BRXf необходима для выключения стандартных паттернов деления.
Важный компонент цитоскелета растительной клетки — кортикальные (то есть находящиеся рядом с цитоплазматической мембраной) микротрубочки, которые играют ключевую роль в ориентации делении. Так называемая препрофазная полоса, образованная микротрубочками, часто совпадает с расположением будущей клеточной стенки, имеющей минимальную площадь, однако не всегда. В 34% случаев отклонение значительно, и при этом флуоресцентные метки к микротрубочкам никогда не локализуются рядом с BASL/BRXf. Если «полумесяц полярности» расположен в том участке мембраны, откуда могла начать строиться самая короткая стенка, — этого не происходит, микротрубочки смещаются.
Именно это наблюдение авторы сочли основным для функционирования BASL/BRXf. Свое предположение они подтвердили с помощью клеточной линии, имеющей мутацию в гене trm678, которая приводит к исчезновению препрофазной полоски. В таких клетках деление может пересекать зону полярного комплекса; таким образом, препрофазная полоска необходима для асимметричного наследования BASL/BRXf. Результат подтвердил и флуоресцентный репортер к MUTE — транскрипционному фактору, определяющему идентичность клеток-предшественников замыкающих клеток устьица. Клетки с мутациями trm678 чаще при делении передавали обеим дочерним клеткам BASL/BRXf, никогда не становились MUTE+. Проростки с мутациями trm678 имели на семядолях меньше устьиц и измененный эпителий.
Влияние BASL/BRXf на препрофазную полоску изучили на клетках-предшественницах устьиц, меченых по микротрубочкам (TMMp). Рядом с полярной областью оказалось очень мало антиклинальных (выгнутых) микротрубочек. Другие данные также подтвердили, что полярность BASL/BRXf необходима для исчезновения микротрубочек у мембраны. При этом формирование поляризованных BASL/BRXf не зависит от микротрубочек (то есть исключена обратная возможность — сборка комплексов в бедных микротрубочками областях мембраны).
Следовательно, структуры растительных клеток, управляющие делением, не «тянут» цитоскелет, как клетки животных, а «отталкивают». «Вместо того, чтобы с помощью цитоскелета сказать: “Делим так!”, растения сказали: “НЕ делим так!”» — говорит ведущий автор статьи Эндрю Мурояма из Калифорнийского университета в Сан-Диего.
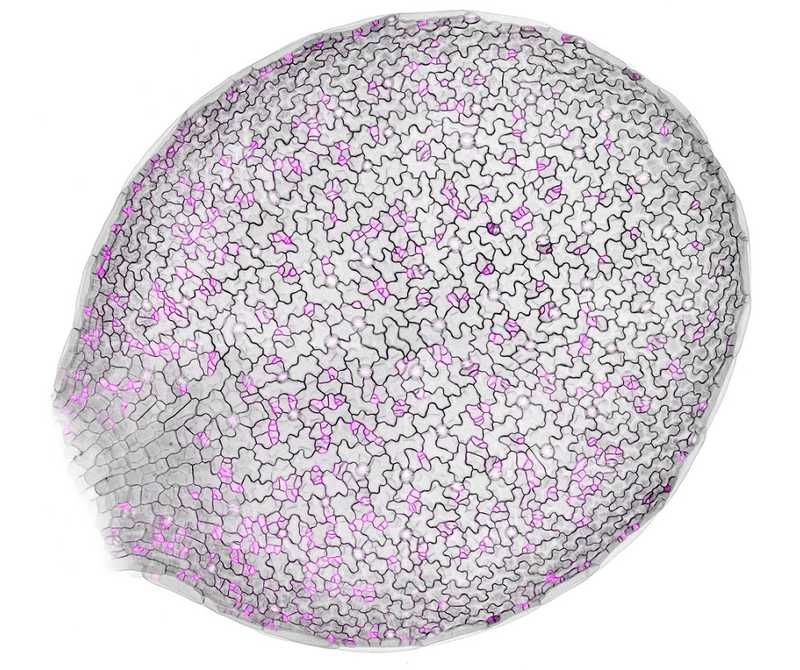 Развивающийся лист Arabidopsis, границы клеток отмечены черным, белок полярности — пурпурным. Credit: Andrew Muroyama
Развивающийся лист Arabidopsis, границы клеток отмечены черным, белок полярности — пурпурным. Credit: Andrew Muroyama
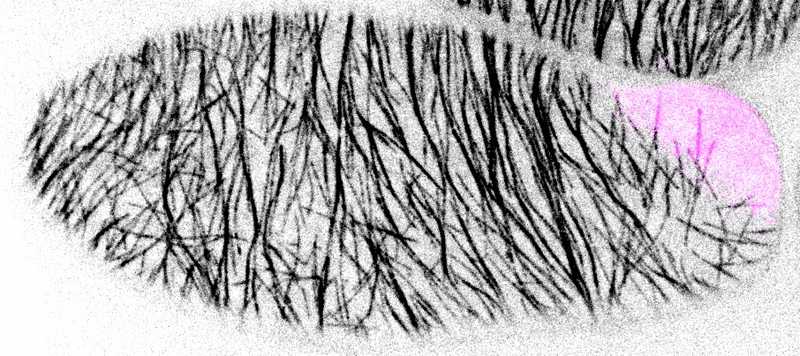 Участок клетки возле мембраны, занятой BASL (пурпурный), обеднен микротрубочками (черный). Credit: Andrew Muroyama
Участок клетки возле мембраны, занятой BASL (пурпурный), обеднен микротрубочками (черный). Credit: Andrew Muroyama
Механизм, за счет которого BASL/BRXf устраняет микротрубочки вокруг себя, установили с помощью гетерологической системы. Репортер микротрубочек TUA5 ввели в клетки с повышенной активностью BASL. Фрагментация микротрубочек важна для их структурной реорганизации, однако рядом с BASL/BRXf она подавлена и преобладают два других эффекта. Во-первых, BASL сильно снижает скорость полимеризации/деполимеризации на плюс-конце микротрубочек. Во-вторых, тот же конец катастрофически разрушается при сближении с BASL.
Таким образом, полярный комплекс обедняет окружающую область мембраны стабильными микротрубочками и тем самым «запрещает» деление, проходящее через данную область, что и становится причиной асимметрии.
Источник
Andrew Muroyama, et al. Cortical polarity ensures its own asymmetric inheritance in the stomatal lineage to pattern the leaf surface // Science (2023), Vol. 381, pp.54-59. Published: 6 July 2023. DOI: 10.1126/science.add6162
Вам будет интересно



 124
124
 0
0