
 Меню
Меню





 Все темы
Все темы
Астроциты участвуют в формировании воспоминаний
За формирование памяти отвечают особые энграммные нейроны, для которых характерна экспрессия транскрипционного фактора c-Fos. Ученые из США выяснили, что их активность регулируется популяцией астроцитов, которые тоже экспрессируют этот фактор транскрипции. Такие ассоциированные с обучением астроциты участвуют в формировании и повторном извлечении воспоминаний.
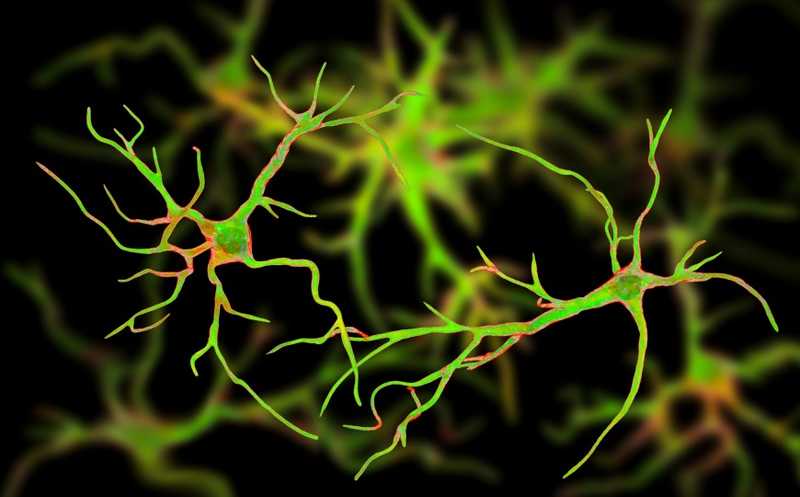
Известно, что за хранение и извлечение воспоминаний отвечают ансамбли нейронов, которые называются энграммами. Астроциты — глиальные клетки мозга — способны регулировать активность нейронов в составе трехчастных синапсов. Ученые из Медицинского колледжа Бэйлора (США) выяснили, что существует особая группа астроцитов, которая также участвует в формировании воспоминаний и регулирует энграммные нейроны.
Исследования памяти в эксперименте проводились на мышах, у которых вырабатывали условный рефлекс в связи с переживанием чувства страха (fear conditioning). Животных помещали в специальные камеры, давали им осмотреться, а затем били их короткими разрядами тока. Через некоторое время мышей возвращали либо в те же камеры, либо в новые. Если животные оказывались в знакомой камере, то замирали от страха, ожидая получить удар током; в новой камере этой реакции не происходило. Таким образом ученые способствовали формированию воспоминаний и их извлечению у животных.
Далее ученые использовали хемогенетические подходы, чтобы специфически активировать нейроны в зубчатой извилине гиппокампа. После этой стимуляции они наблюдали экспрессию c-Fos в зубчатой извилине и в области CA1 гиппокампа — c-Fos относится к немедленно-ранним генам (immediate-early genes), которые начинают экспрессироваться первыми при активации клеток мозга. В нейронах экспрессия c-Fos обеспечивает синаптическую пластичность и формирование памяти. После экспериментов с условно-рефлекторным замиранием, т.е. после обучения животных, экспрессия c-Fos активировалась в астроцитах, причем пик экспрессии достигался через 1,5 ч после такого опыта.
Если же ген Fos нокаутировали в астроцитах гиппокампа с помощью Cre-lox рекомбинации, долгосрочная потенциация в гиппокампе ослабевала — это подтвердило, что активация астроцитов необходима для формирования памяти. Нокаут c-Fos отразился и на поведении мышей в поведенческих экспериментах: у них не формировался условный рефлекс от ударов током.
Популяцию астроцитов, в которых экспрессируется c-Fos в ответ на активность нейронов в гиппокампе, ученые назвали ассоциированными с обучением — learning-associated astrocytes (LAA). Они получили репортерную линию мышей Aldh1l1-CreER;Rosa-CAG-FSF-tdTomato, в которой используется две сайт-специфичные рекомбиназы — Cre и Flp — для специфической экспрессии флуоресцентного белка tdTomato в LAA. Действительно, после проведения экспериментов с условно-рефлекторным замиранием в гиппокампе стали появляться астроциты, экспрессирующие tdTomato, причем в них повышалась активность кальциевого сигналинга.
Пометив энграммные нейроны и LAA — обе группы клеток экспрессируют c-Fos при запоминании животными событий — разными флуоресцентными метками, ученые выяснили, что астроциты, связанные с памятью, располагаются рядом с энграммными нейронами и с их синапсами. Исследователи проверили, регулируют ли астроциты их активность. Для этого они специфически активировали LAA, что приводило к возникновению долговременной потенциации в энграммных нейронах. В них также повышалась частота возбуждающих постсинаптических токов. В еще одном эксперименте ученые пометили энграммные нейроны и LAA, которые активировались при формировании у мышей чувства страха. Через неделю эти LAA активировали, а затем поместили мышей в новую камеру. Несмотря на то, что эта камера была им незнакома, из-за активации LAA все равно активировались энграммные нейроны, хранящие воспоминания о том, что происходило в первой камере.
Оказалось, что LAA нужны не только для формирования воспоминаний, но и для их извлечения. При активации LAA, связанных с первой камерой, мыши замирали, находясь в новой камере, хотя у них не было причин ожидать получить удар током и в ней из-за смены обстановки. Этот феномен сохранялся в течение трех недель после первого эксперимента в старой камере.
Транскриптомный анализ показал, что для извлечения воспоминаний необходим транскрипционный фактор NFIA, роль которого в процессах, связанных с памятью, обсуждалась и ранее. Исследователи селективно нокаутировали NFIA в LAA, а потом проверили, как поведут себя мыши при повторном помещении в тот же контекст, где у проводилось обуславливание страха. У таких мышей не формировался условный рефлекс: они не замирали от страха. Тот же эффект до этого наблюдался при нокауте c-Fos. Вместе с тем этот эффект не был замечен в других условиях: когда мышей проверяли на способность узнавать новые места. Следовательно, LAA, вероятно, необходимы для консолидации и извлечения конкретных воспоминаний, связанных с теми или иными событиями.
Тем не менее, извлечение воспоминаний зависит от активности именно энграммных нейронов, а не от LAA, хотя они и принимают участие в этом процессе. Ученые сформировали у мышей условный рефлекс замирания, пометили энграммные нейроны и LAA, а затем нокаутировали из LAA транскрипционный фактор NFIA. Через три недели они повторно активировали энграммные нейроны, а потом поместили мышей в новую камеру, где мыши начали замирать от страха, вспоминая об ударах током в предыдущей камере. Это показало, что активность энграммных нейронов необходима и достаточна для извлечения воспоминаний, несмотря на то что LAA тоже участвуют в этом процессе.
Источник
Вам будет интересно
 177
177
 0
0


 118
0
118
0








