Меню
Меню





 Все темы
Все темы
Присоединение моноаминовых меток к гистонам влияет на циркадные ритмы
Посттрансляционные модификации гистонов играют ключевую роль в эпигенетической регуляции, влияя на архитектуру хроматина и доступность генов. Ранее исследователи из США показали, что моноамины серотонин и дофамин присоединяются к гистону H3, регулируя экспрессию генов. В новой работе ученые открыли еще один вид моноаминовых модификаций H3 — присоединение гистамина. Все три вида меток присоединяет и удаляет фермент TG2, и этот процесс важен для регуляции циркадных ритмов у млекопитающих.
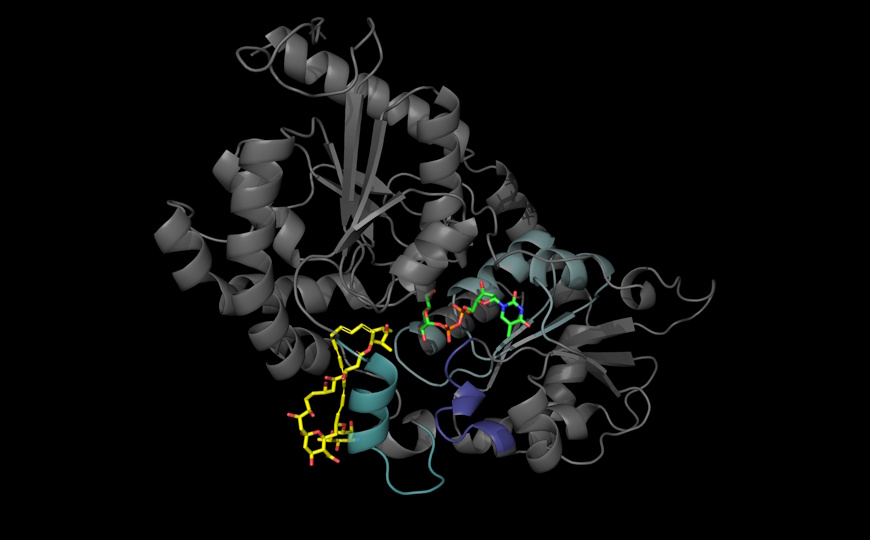
Модификации гистонов — одни из ключевых эпигенетических регуляторных механизмов. Хорошо изученные модификации, такие как метилирование, ацетилирование и убиквитинирование, динамически регулируются сайтспецифическими ферментами — писателями, присоединяющими метки, и стирателями, удаляющими их (writer and eraser). Присоединенные группы распознаются белками-считывателями (reader). Кроме того, с субстратными белками могут напрямую реагировать многочисленные низкомолекулярные метаболиты. Все это влияет на архитектуру хроматина и доступность генов для транскрипции, следовательно, и на судьбу клеток.
Авторы новой статьи в Nature ранее сообщили об открытии нового класса посттрансляционных модификаций гистонов — моноаминилирования. Моноамины, такие как серотонин (подробнее на PCR.NEWS) и дофамин, ковалентно связываются с остатком глутамина в положении 5 гистона H3 (это называется серотонилированием и дофаминилированием, или H3Q5ser и H3Q5dop). Серотонилирование гистонов, триметилированных по лизину в положении 4 (H3K4me3) усиливает экспрессию и ослабляет деметилирование H3 за счет ингибирования деметилаз KDM5 и LSD1. Дофаминилирование в нейронах влияет на транскрипционные программы таким образом, что возрастает риск рецидива при отказе от наркотиков. Реакции моноаминирования катализируются Ca2+-зависимым ферментом трансглутаминазой 2 (TG2).
Совместные исследования сотрудников Медицинского центра Маунт-Синай и Мемориального онкологического центра Слоуна–Кеттеринга в Нью-Йорке показали, как моноаминные нейротрансмиттеры участвуют в регуляции активности мозга. Они исследовали динамику моноаминирования гистона H3 и определили, что TG2 может действовать как писатель, стиратель и модулятор моноаминированных фрагментов на гистоне H3.
TG2 способен присоединять любой из моноаминовых кофакторов, в зависимости от их концентрации в нуклеоплазме. Более того, когда TG2 выступает в качестве стирателя, в отсутствие других моноаминов он превращает в месте модификации глутамин в глутамат, что, по предположению авторов, пагубно влияет на функциональную активность нейрона.
Авторы идентифицировали третий класс моноаминилирования H3 — гистаминилирование по глутамину в пятом положении (H3Q5his), которое также выполняет фермент TG2. Гистаминирование Н3 препятствует его метилированию по соседнему лизину ферментом MLL1 и снижает связывание белка WDR5, который функционирует как считыватель серотониновой метки, способствующей экспрессии генов. Ранее было показано, что гистамин в неонатальном мозге грызунов в основном локализован в ядрах клеток.
Таким образом, TG2 присоединяет, удаляет и обменивает гистаминовые, серотониновые и дофаминовые метки на Н3, и это первое сообщение об эпигенетическом регуляторе с такой активностью.
Заднее туберомаммиллярное ядро гипоталамуса (TMN) — единственная область мозга, которая содержит нейроны, экспрессирующие гистидиндекарбоксилазу, фермент, который катализирует образование гистамина из гистидина. Эта область в основном состоит из гистаминергических нейронов и участвует в различных биологических функциях, от контроля возбуждения до поддержания циклов сна-бодрствования и энергетического баланса. Более того, к TMN сходятся проекции многих отделов мозга, включая ствол мозга, серотонинергические структуры, которые деполяризуют нейроны TMN; другие моноаминергические структуры мозга, такие как голубое пятно (норадренергическое ядро) и черная субстанция/VTA (дофаминергическое ядро), посылают только небольшое количество волокон в TMN. Поэтому авторы выбрали TMN для изучения гистаминилирования гистонов и его возможного влияния на циркадные колебания экспрессии генов.
Действительно, в гистонах TMN гистаминовая метка встречается значительно чаще, чем в других ядрах мозга. РНК-секвенирование образцов TMN подтвердило, что многие гены, участвующие в регуляции циркадного цикла (такие как Arntl, Dbp, Per1/2) демонстрируют циклические колебания экспрессии, связанные с активностью ключевого регулятора циркадных ритмов CLOCK.
Далее авторы исследовали влияние циркадных ритмов и цикла сна и бодрствования мышей на моноаминилирование гистонов. Уровень гистаминовой метки значительно колебался в течение суточного цикла, достигая пика во время активной фазы и снижаясь во время неактивной. На эти изменения не влияли суточные колебания экспрессии TG2, гистидиндекарбоксилазы и общие уровни моноаминов в TMN. Уровень серотонилирования гистона Н3 также изменялся в течение суток, и он сильнее снижался к концу активной фазы, чем уровень гистаминилирования.
Иными словами, паттерны моноаминилирования гистонов коррелируют с ритмическими паттернами экспрессии генов, которые регулируются факторами транскрипции CLOCK–BMAL1. Интересно, что на уровни моноаминилирования гистонов в TMN оказывал влияние снотворный препарат золпидем. Когда мышь получала снотворное (искусственно вызванная неактивная фаза), уровни H3Q5his, H3Q5ser и WDR5 снижались по всему геному и особенно в циркадных генах, которые регулируются факторами транскрипции CLOCK и BMAL1.
Чтобы доказать причинно-следственную связь между моноаминилированием гистона Н3 и циркадными ритмами, авторы получили мышей, у которых в нейронах TMN экспрессировался Н3 без глутаминового остатка в пятом положении. У таких мышей нарушались суточные ритмы экспрессии генов и двигательной активности.
 Исчезновение серотониновой метки ослабляет связывание WDR5 с гистоном, в итоге это приводит к диссоциации BMAL1 и CLOCK и началу неактивной фазы суточного цикла. Credit: Nature 2025. DOI:
10.1038/s41586-024-08371-3
Исчезновение серотониновой метки ослабляет связывание WDR5 с гистоном, в итоге это приводит к диссоциации BMAL1 и CLOCK и началу неактивной фазы суточного цикла. Credit: Nature 2025. DOI:
10.1038/s41586-024-08371-3
«Наши результаты показывают, что химические моноаминовые нейротрансмиттеры влияют на внутренние часы мозга ранее неизвестным способом, — говорит руководитель работы Иэн Мейз. — А именно, моноамины могут напрямую модифицировать гистоны, которые, в свою очередь, регулируют паттерны экспрессии циркадных генов мозга, нейронную пластичность и активность сна или бодрствования». Моноаминилирование способно влиять на пластичность нейронов, а также участвовать в патогенезе различных заболеваний нервной системы.
Источник
Zheng, Q., et al. Bidirectional histone monoaminylation dynamics regulate neural rhythmicity // Nature 2025. DOI: 10.1038/s41586-024-08371-3
Цитата по пресс-релизу

Вам будет интересно

 231
231
 0
0


 624
0
624
0
 1281
0
1281
0