Меню
Меню





 Все темы
Все темы
Роль клеток-канделябров в зрительной коре мышей установили с помощью виртуальной реальности
Клетки-канделябры, присутствующие в первичной зрительной коре, до сих пор оставались слабо изученными. Теперь же ученые из Нидерландов проанализировали их роль в обработке визуальной информации. Для этого они помещали мышей в виртуальный коридор, стены которого были покрыты текстурой гауссовского шума с наложенными на него визуальными эффектами. Оказалось, что клетки-канделябры модулируют активность пирамидных нейронов зрительной коры в ответ на неожиданные стимулы — например, на несоответствие визуальных и моторных сигналов.
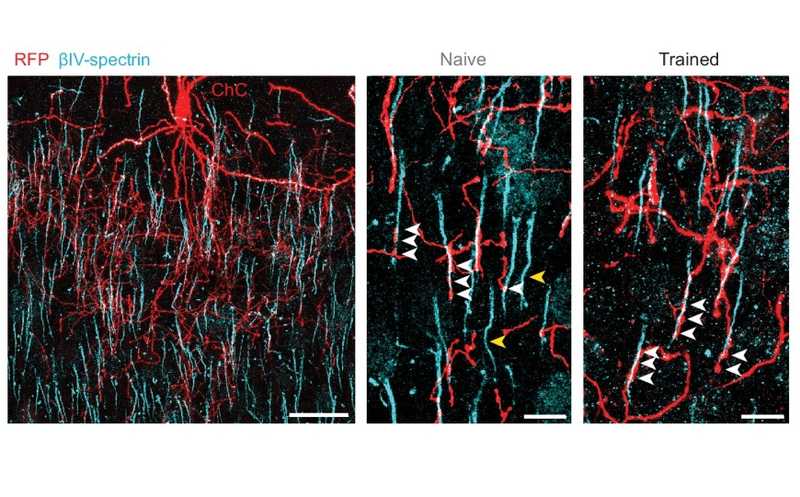
Конфокальная микроскопия слоя 2/3 зрительной коры: клетки-канделябры отмечены красным, начальные сегменты аксонов пирамидных клеток — голубым. Стрелками отмечены точки колокализации клеток канделябров и начальных сегментов аксонов у наивных и обученных мышей.
Credit:
eLife (2024). DOI:
10.7554/eLife.91153.3 |
CC BY
Тормозные интернейроны в неокортексе очень разнообразны. Аксо-аксональные клетки-канделябры (chandelier cells, ChC) остаются мало изученными, несмотря на то, что большинство подгрупп кортикальных интернейронов в первичной зрительной коре (V1) хорошо охарактеризованы. Эти клетки отличаются от других типов интернейронов тем, что они иннервируют исключительно пирамидальные клетки (PyC) в начальном сегменте аксона — месте, где генерируются потенциалы действия (ПД). Такая анатомическая организация привела к идее, что ChC могут осуществлять мощный контроль над генерацией ПД. Недавнее открытие того, что рецептор вазоактивного кишечного пептида 2 (vasoactive intestinal peptide receptor 2, Vipr2) служит маркером кортикальных ChC, и создание линии мышей Vipr2-Cre позволили провести тщательный анализ этого малоизученного типа клеток.
Специалисты из Нидерландов проанализировали клетки-канделябры в поверхностном слое 2/3 (L2/3) первичной зрительной коры, чтобы прояснить, какую роль они играют в обработке визуальной информации. Они показали, что ChC преимущественно реагируют на возбуждение, связанное с передвижением или неожиданными стимулами, и действуют, слабо модулируя активность и/или пластичность пирамидальных клеток в зрительной коре. Результаты работы они опубликовали в журнале eLife.
Вначале ученые идентифицировали источники синаптического сигнала для клеток-канделябров в зрительной коре. Чтобы визуализировать эти клетки в слое L2/3, они использовали мышей Vipr2-Cre, у которых в кортикальных ChC избирательно экспрессируется рекомбиназа Cre. В поверхностный слой зрительной коры животных ученые ввели Cre-зависимые аденоассоциированные вирусные векторы (AAV), несущие флуоресцентный белок и гликопротеин вируса бешенства. Спустя четыре недели мышам инъецировали вирус бешенства, лишенный гликопротеина, после чего отслеживали распространение флуоресцентной метки. С помощью этого подхода ученые выяснили, что ChC получали сигналы из различных областей таламуса и коры (например, дорсальной части латерального коленчатого ядра и латерального заднего ядра таламуса, а также ретроспленальной и соматосенсорной коры). Однако наиболее распространенные пресинаптические партнеры ChC были обнаружены локально в слое L5 и в меньшей степени в слоях L1–4 зрительной коры. Меченые нейроны слоя L5 имели пирамидальные тела клеток и дендритные шипики — это указывает на то, что синапсы с ними являются возбуждающими.
Затем ученые проанализировали поведение ChC и PyC слоя L2/3 in vivo с использованием двухфотонной визуализации кальция у бодрствующих животных. Они оценили активность этих клеток во время спонтанного поведения, отслеживая скорость бега на беговом колесе и размер зрачков, когда мыши смотрели на однородный серый экран. В качестве области обзора авторы выбрали границу между слоями L1 и L2/3, где расположены поверхностные ChC. Клетки-канделябры были наиболее активны в состояниях сильного возбуждения, в большей степени, чем PyC. Кроме того, проявления активности ChC в одном и том же поле зрения сильно коррелировали друг с другом, что позволяет предположить, что они распространяют синхронизированный сигнал во время сильного возбуждения. Однако клетки-канделябры оказались слабо избирательны к зрительной информации.
В дальнейшей работе ученые использовали сконструированную ими установку виртуальной реальности. Мыши, бегущей в колесе, демонстрировали изображение коридора. Его левая половина отображалась на мониторе, расположенном под углом 45 градусов и просматриваемом через зеркало, что создавало иллюзию симметричного туннеля. Коридор имел длину 100 см, а его стены были покрыты черно-белой текстурой гауссовского шума с наложенными поверх визуальными сигналами. Эти сигналы представляли собой три решетки и три изображения шахматной доски, расположенные на расстоянии 11 см друг от друга. После пробега по визуальной части коридора мышам показывали серый экран, через секунду после начала демонстрации которого подавали звуковой сигнал, а затем через две секунды давали 5 мкл воды в качестве вознаграждения.
Ученые выяснили, что в начале зрительного участка туннеля клетки-канделябры были высокоактивны, но подавлялись, когда мыши видели зрительные стимулы. ChC в основном управлялись локомоцией и демонстрировали сильную реакцию на зрительно-моторное несоответствие в модельной ситуации, в которой исследователи ненадолго (0,5 с) прерывали зрительный поток, пока мышь бежала. Такое несоответствие стимулов подавляло активность ChC.
В целом проведенные эксперименты показали, что ChC преимущественно реагировали на локомоцию и зрительно-моторное несоответствие, когда зрительный поток прекращался во время бега. В то же время их активность подавлялась зрительными стимулами или началом зрительного потока в то время, когда мыши были неподвижны.
Кроме того, повторное воздействие виртуального туннеля на мышей сопровождалось снижением реакций клеток-канделябров. Эксперименты с виртуальным туннелем продемонстрировали пластичность длины начального сегмента аксона пирамидальных клеток и характера их иннервации клетками-канделябрами. Также ученые обнаружили, что ChC оказывали лишь слабое ингибирующее влияние на PyC, затрагивая небольшую часть клеток.
Результаты проведенных экспериментов позволяют предположить, что клетки-канделябры передают сигнал, связанный с возбуждением, пирамидальным клеткам слоя 2/3, ограниченно модулируя их активность и/или пластичность начальных сегментов их аксонов во время поведенчески значимых событий.
Вкусные и невкусные запахи обрабатываются у саранчи разными нейронами
Источник
Seignette K. et al. Experience shapes chandelier cell function and structure in the visual cortex (2024). // eLife 12: RP91153. DOI: 10.7554/eLife.91153.3





 0
0