Меню
Меню





 Все темы
Все темы
Зачем трипаносоме 2,5 тысячи поверхностных антигенов?
Для ухода от иммунного ответа многие патогены прибегают к антигенной изменчивости. Например, Trypanosoma brucei — возбудитель сонной болезни, — паразитируя в организме млекопитающих, регулярно переключает экспрессию поверхностных гликопротеинов, избегая узнавания антителами. Авторы статьи в Nature адаптировали метод секвенирования РНК единичных клеток Smart-seq, чтобы установить механизмы такого переключения в клетках трипаносомы. Они обнаружили два ключевых фактора, которые регулируют активацию тех или иных поверхностных гликопротеинов.
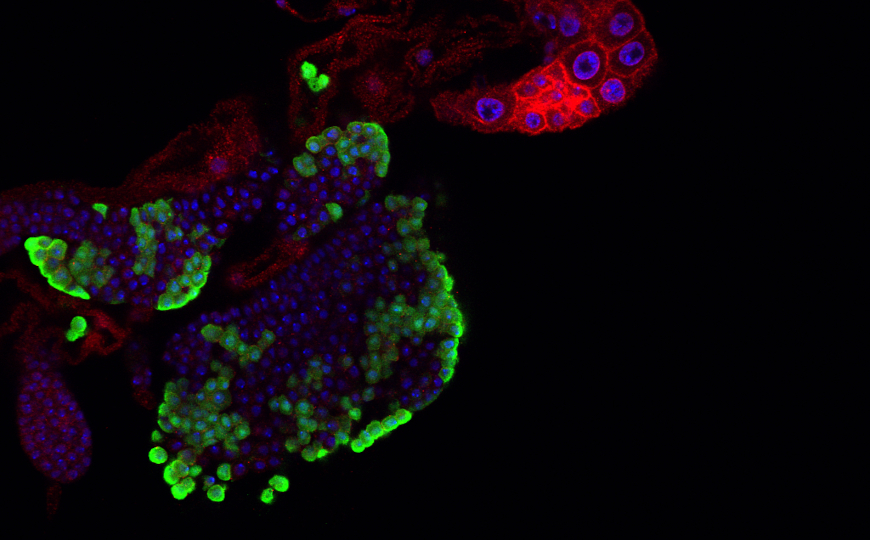
Антигенная изменчивость — это стратегия уклонения от иммунитета, используемая многими патогенами. Она опирается на регулярные переключения паттернов экспрессии поверхностных антигенов. Для изучения этого процесса необходимо отслеживать изменения транскриптома и возможные геномные перестройки в отдельных клетках патогена, однако метода недоставало. Авторы статьи в Nature разработали высокочувствительный метод секвенирования РНК для анализа изменений, происходящих в отдельной клетке паразита Trypanosoma brucei.
Наружный слой оболочки трипаносомы формируют варианты поверхностных гликопротеинов (VSG). Избежать распознавания антителами и выведения из кровотока этому паразиту удается за счет антигенной изменчивости — в геноме трипаносомы содержится около 2 500 генов VSG, между которыми она может переключаться. Большинство генов поверхностных гликопротеинов хранятся в молчащих субтеломерных массивов, однако еще часть кодируется в высокоспециализированных минихромосомах.
Уже было показано, что двуцепочечные разрывы в области активно транскрибируемых VSG переключают паттерны их экспрессии. Чтобы исследовать механизм отбора, авторы создали систему на основе Cas9 и с ее помощью индуцировали двуцепочечные разрывы в различных местах области активной экспрессии. Они снижали экспрессию целевого VSG — к четвертому дню она была утрачена у более чем 94% популяции.
Секвенирование тотальной РНК не позволяет отличить моногенную экспрессию VSG от мультигенной и не дает информации о механизмах или месте переключения. Поэтому ученые адаптировали для анализа единичных клеток T. brucei последнюю версию протокола Smart-seq. Они модифицировали стандартные олиго(dT)-праймеры Smart-seq3xpress, опустили этап переключения матрицы и оптимизировали концентрацию транспозазы, чтобы учесть малые количества РНК в клетках трипаносомы по сравнению с клетками млекопитающих.
Авторы сравнили результаты, полученные их методом, с лучшим из доступных наборов данных от 10X Genomics. При одинаковой глубине секвенирования SL-Smart-seq3xpress оказался намного чувствительнее — он выявил в среднем 2 876 генов на клетку против 1 052 в уже опубликованных данных.
Затем ученые применили разработанный подход для анализа механизма переключения VSG. Они выбрали в качестве мишени VSG-2 — активный вариант в большинстве лабораторных изолятов трипаносомы — и отслеживали экспрессию VSG в отдельных клетках до индукции двуцепочечных разрывов и в разные моменты после. Во всех случаях разрывы внутри VSG-2 или чуть ближе к 5’-концу активировали примыкающий к теломере VSG.
Однако зачем паразиту иметь в запасе более 2 500 генов VSG, если активируются только прилегающие к теломерам? Углубившись в этот вопрос, ученые обратили внимание на уникальность последовательности VSG-2 по сравнению с другими VSG, и решили проверить, влияла ли низкая гомология на механизм. Они получили клетки трипаносомы с активным VSG-8 или VSG-11, последовательность которых куда ближе к остальным VSG. Оказалось, что двуцепочечные разрывы в таких клетках репарировались путем сегментной конверсии генов, а не при помощи индуцированной разрывом репликации, как в случае с VSG-2. По-видимому, T. brucei сначала ищет гомологичную последовательность VSG, чтобы использовать ее в качестве шаблона для репарации, и только в случае неуспеха переключается на репликацию, индуцированную повреждением ДНК.
Разработанный метод, таким образом, позволил установить, что ключевыми факторами, определяющими иерархию активации VSG, служат доступность гомологичной ДНК для залечивания двуцепочечных разрывов и расположение VSG в геноме. При наличии подходящей матрицы репарация происходила путем сегментной перестройки, приводя к образованию новых антигенов. В противном случае активировался примыкающий к теломере VSG. Авторы надеются, что их результаты углубят понимание эволюции патогенов и в дальнейшем лягут в основу методов борьбы с ними.
Малярийного плазмодия заставили экспрессировать лишь один вариант белка PfEMP1
Источник

Вам будет интересно
 116
116
 0
0