Меню
Меню





 Все темы
Все темы
Гормон мозга защищает кости кормящей матери от остеопороза
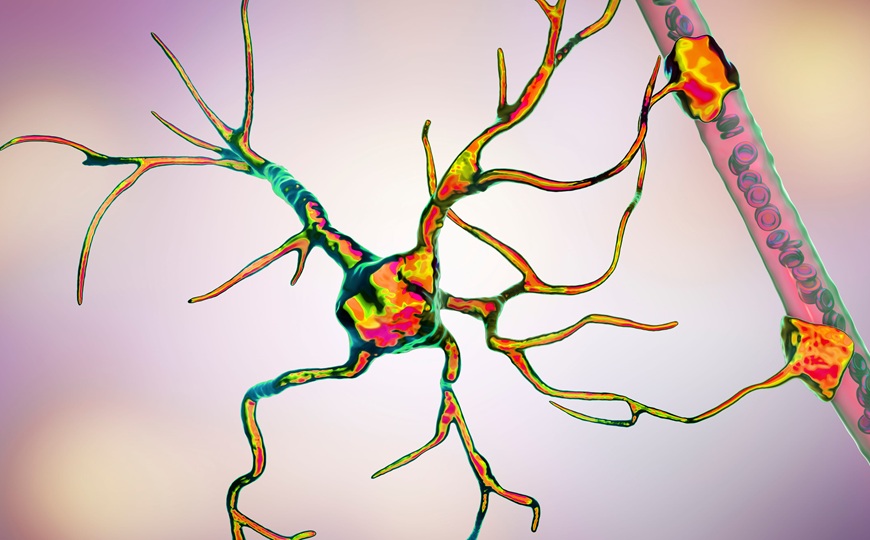
В организме кормящей матери одновременно происходит передача кальция из скелета матери детенышам и активация процессов, укрепляющих кости. Исследователи из США показали, что за регуляцию этих процессов отвечает гормон CCN3, экспрессируемый в аркуатном ядре гипоталамуса. Гель, содержащий CCN3, может ускорять заживление переломов.

Во время грудного вскармливания кальций, в котором нуждается новорожденный, поступает в молоко, а его содержание в скелете матери уменьшается. Это приводит к значительному снижению костной массы — до 30% у грызунов и 10% у человека. У самок млекопитающих эстрадиол действует как анаболический гормон в отношении костной ткани, то есть способствует сохранению костной массы (поэтому после менопаузы у женщины могут появиться признаки остеопороза). Однако во время лактации уровень эстрадиола, поднявшийся на поздних стадиях беременности, резко падает, а вымыванию кальция из костей способствует PTHrP (белок, родственный паратиреоидному гормону). Очевидно, должен существовать компенсаторный механизм, предотвращающий критическое ослабление скелета при вскармливании потомства. Ученые из Калифорнийского университета в Сан-Франциско и Дейвисе показали, что за наращивание костной массы во время лактации отвечает гормон CCN3 (communication network factor 3). Этот гормон также имеет высокий потенциал для заращивания переломов.
Скелетные перестройки во время лактации ассоциированы с понижением уровня эстрогена. В предыдущей работе авторы показали, что делеция рецепторов эстрогена в аркуатном ядре гипоталамуса (один из центров мозга, отвечающих за энергетический метаболизм и поддержание гомеостаза организма) у мышей приводит к значительному повышению костной массы уже к возрасту четырех недель. В данном исследовании ученые хирургически объединяли кровеносные системы самок мутантных мышей и мышей дикого типа (метод, называемый парабиозом). В таких парах наблюдалось повышение костной массы, характерное для мутантного фенотипа. В парах из двух животных дикого типа этого не происходило. Схожий эффект обнаруживался при пересадке бедренной кости от мутантных самок животным дикого типа обоих полов. Эти результаты указывали на то, что регуляция повышенного костного роста осуществляется гуморальным (циркулирующим в крови) фактором, вырабатываемым только у самок, но работающим и у самок, и у самцов.
Затем мутантным животным пересадили под почечную капуслу остеохондральные скелетные стволовые клетки от мышей дикого типа. У мышей в месте пересадки образовалась костная ткань, значительно более плотная, чем при пересадке таких же клеток животным дикого типа. Кроме того, у мутантных мышей образование минерализованных структур наблюдалось и при пересадке стволовых клеток в непосредственной близости от аркуатного ядра, при этом у мышей дикого типа таких образований не появлялось. Это подтверждало, что фактор, стимулирующий рост костной ткани, вероятнее всего, синтезируется в аркуатном ядре, оставалось только его идентифицировать.
Для этого исследователи проанализировали изменения в экспрессии генов в данной части мозга у мутантных мышей, получавших богатую жиром диету — такой режим питания влияет на активность нейронов аркуатного ядра. Диета действительно обращала вспять повышение костной массы у мутантных мышей. При этом изменялись уровни экспрессии четырех генов. Три из этих генов активно экспрессировались в аркуатном ядре в возрасте до четырех недель, а среди них только один — Ccn3 — успешно повышал уровень минерализации в культурах скелетных стволовых клеток.
Дальнейшие опыты на более сложных моделях подтвердили, что CCN3 способствует костному росту. Гормон повышал костную массу как в ex vivo препаратах бедренной кости, так и in vivo при введении внутривенно мышам дикого типа. Более того, наложение гидрогеля с CCN3 на место перелома у пожилых мышей (у которых такие повреждения заживают плохо) приводила к образованию плотной и крепкой костной ткани на месте перелома. Один из руководителей работы Томас Эмбрози отмечает, что им не удавалось добиться подобного результата никакими другими методами.
Наконец, авторы описали роль CCN3 в метаболизме кальция во время лактации. Уже через неделю после деторождения уровни CCN3 в мозгу самок сравнимы с теми, что наблюдаются у мутантных животных. Если самка переставала кормить, уровень гормона снижался. Нокаут CCN3 перед беременностью приводил к более выраженному снижению костной массы самки после выкармливания потомства. А когда CCN3-дефицитные самки получали диету с низким содержанием кальция, смертность среди потомства существенно повышалась.
Таким образом, авторы установили роль гормона CCN3 в регуляции кальциевого обмена во время вскармливания потомства, а также продемонстрировали его потенциал как средства для лечения переломов. Они планируют проведение дальнейших исследований для установления молекулярных механизмов действия CCN3.
«Потеря костной массы происходит не только у женщин в постменопаузе, она часто встречается у выживших после рака груди, которые принимают определенные гормональные блокаторы; у молодых, хорошо тренированных элитных спортсменок; и у пожилых мужчин, чья относительная выживаемость ниже, чем у женщин после перелома шейки бедра, — сказала соруководительница работы Холли Ингрэм. — Было бы невероятно захватывающе, если бы CCN3 мог увеличить костную массу во всех этих сценариях».
Риск перелома бедренной кости можно предсказать по протеому крови
Источник
Babey, M.E. et al. A maternal brain hormone that builds bone. // Nature, published July 10, 2024, DOI: 10.1038/s41586-024-07634-3
Цитаты по пресс-релизу
Вам будет интересно

 45
45
 0
0


 37
0
37
0