Меню
Меню





 Все темы
Все темы
Как клетка преодолевает зависимость от глутамина
Аминокислота глутамин не является незаменимой, но в быстро пролиферирующих клетках (например, раковых) может развиться зависимость от поступления глутамина извне. Однако такие клетки выживают и при недостатке глутамина. Исследователи из США и Швейцарии показали, что для выживания им необходим дополнительный источник углерода, а не азота, например, пируват. Скрининг продемонстрировал, что биотинилирование пируваткарбоксилазы является лимитирующим фактором для роста клеток, зависимых от глутамина. А опухолевый супрессор FBXW7 способствует пролиферации клеток в отсутствие глутамина, поддерживая экспрессию пируваткарбоксилазы. Таким образом, витамин биотин и FBXW7 необходимы для преодоления зависимости от глутамина.
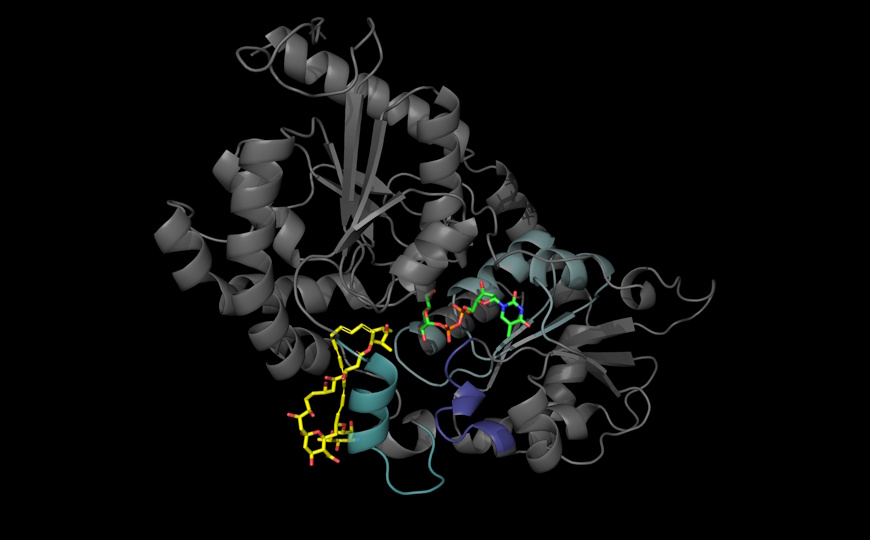
Глутамин — наиболее распространенная аминокислота в плазме крови и одна из наиболее активно используемых клетками в культуре. Она не только входит в состав белков, но и участвует в образовании АТФ через цикл трикарбоновых кислот, а также в метаболизме азота. Глутамин не является незаменимой аминокислотой, но в клетках млекопитающих может развиться зависимость от поступления глутамина извне. Считается, что такая зависимость от глутамина появляется, когда скорость пролиферации слишком велика, и синтезируемого внутри клетки глутамина не хватает. Неясно, с чем это может быть связано, — с недостатком углерода или азота. Зависимость от глутамина характерна для некоторых раков. Теоретически этим можно воспользоваться для борьбы с ними. Исследователи из США и Швейцарии выяснили, какие нутриенты и сигнальные пути необходимы для выживания клеток при недостатке глутамина.
Сначала авторы рассмотрели метаболические реакции, в которые вовлечен глутамин и образованный из глутамина глутамат у человека. Среди 20 420 белоккодирующих генов 43 использовали глутамин, а 127 — глутамат. Эти белки в основном кодируют метаболические ферменты, транспортеры и рецепторы глутамата, и среди них 28 белков, связанных с глутамином, и 68 белков, связанных с глутаматом, экспрессируются в клетках миелогенной лейкемии человека K562. Опыты с этими клетками показали, что причина глутаминовой зависимости — в недостатке углерода, а не азота.
При недостатке глутамина снижается количество всех промежуточных соединений цикла трикарбоновых кислот, а также аминокислот, таких как глутамат, аспартат и аспарагин. Уровни метаболитов, связанных с гликолизом, синтезом нуклеотидов и других аминокислот, а также с пентозофосфатным путем, не менялись значимо. Авторы проверили, может ли другой источник углерода восполнить возникший недостаток. Добавление пирувата действительно восстановило первоначальные уровни промежуточных метаболитов цикла трикарбоновых кислот, глутамата, аспартата и аспарагина.
Недостаток глутамина также увеличивал соотношение NADH/NAD+, чего можно было ожидать при сниженной активности цикла трикарбоновых кислот. Пируват восстанавливал баланс. Более того, в отсутствие глутамина пируват повышал скорость пролиферации клеток K562. То же было показано в сфероидах из клеток HEK239T.
С помощью CRISPR-скрининга авторы подтвердили, что следующие ферменты участвуют в выживании клеток в отсутствие глутамина: митохондриальный компонент малат-аспартатного челночного механизма (GOT2, MDH2 и SLC25A13), пируваткарбоксилаза (катализирует обратимую реакцию образования оксалоацетата из пирувата), аспарагинсинтаза, чувствительный к аминокислотам компонент интегрированного стрессового ответа (GCN1 и GCN2) и система обмена цистеина и глутамата плазматической мембраны Xc- (SLC3A2 и SLC7A11). Также они выявили гены и сигнальные пути, роль которых в выживании при недостатке глутамина ранее не была показана: гены, связанные с биотином (SLC5A6 и HLCS), цитозольный белок, участвующий в трансляции белков, направляемых в митохондрии (CLUH); гены, связанные с аутофагией (ATG), а также пути, связанные с убиквитинированием (GID4, MAEA, WDR26 и YPEL5) и компонент распознавания субстрата E3-лигазы Skp1-Cullin-F-box (FBXW7). Полученные результаты подтвердили генетическими и фармакологическими путями.
Биотин, также известный как витамин B7, является субстратом для биотинилирования. Скрининг показал, что транспортер биотина в плазматической мембране (SLC5A6) и фермент, конъюгирующий биотин (HLCS), необходимы для пролиферации клеток в отсутствие глутамина. Биотинилирование пируваткарбоксилазы является лимитирующим фактором для роста клеток, зависимых от глутамина.
Клетки с дефицитом опухолевого супрессора FBXW7 пролиферировали в среде, богатой глутамином, однако в отсутствие глутамина росли медленнее, несмотря на добавление пирувата. Таким образом, FBXW7 способствует пролиферации клеток в отсутствие глутамина, поддерживая экспрессию пируваткарбоксилазы. Это удивительно, так как основная функция FBXW7 заключается в деградации белков. Оказывается, что при недостатке FBXW7 накапливается протоонкоген c-MYC, в результате репрессоры транскрипции связываются с геномной ДНК и привлекают гистоновые деацетилазы к лизину 27 гистона 3 (H3K27) в промоторе пируваткарбоксилазы, что снижает экспрессию этого фермента. FBXW7 — один из наиболее часто мутирующих генов при раковых заболеваниях.
Кислая среда опухоли заставляет раковые клетки отказываться от гликолиза и активировать митохондрии
Источник:
Miriam Lisci, et al. Functional nutrient-genetic profiling reveals biotin and FBXW7 are essential to bypass glutamine addiction // Molecular Cell (2026), published 25 February 2026, DOI: 10.1016/j.molcel.2026.02.002

Вам будет интересно

 93
93
 0
0


 92
0
92
0
 112
0
112
0
 132
0
132
0