Меню
Меню





 Все темы
Все темы
Как митохондрии участвуют в перепрограммировании клеток
Ученые установили, что пластичность эпителиальных клеток, позволяющая им дедифференцироваться и возвращаться к пролиферации, связана с активностью митохондрий. Ключевым участником описанного пути является киназа Sgk1, активность которой индуцируется активными формами кислорода.
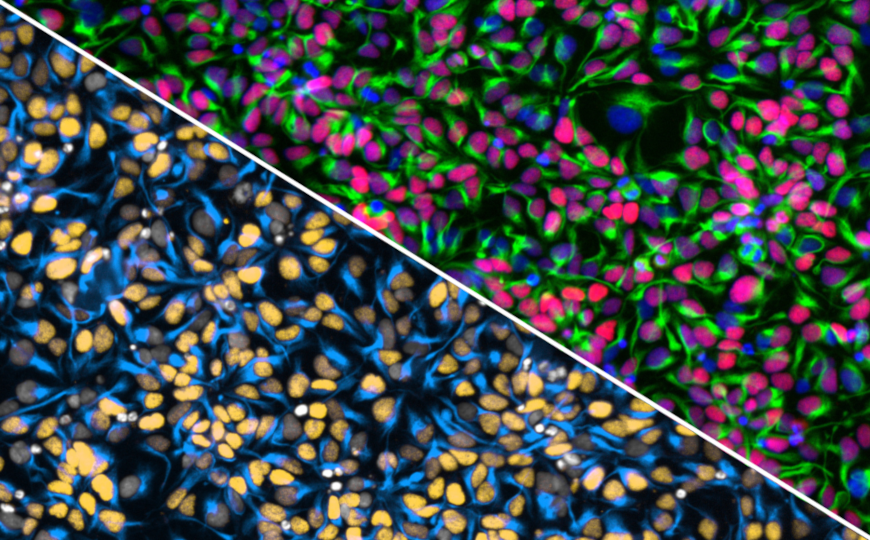
Клеточная пластичность играет ключевую роль в поддержании тканевого гомеостаза, регенерации и онкогенезе. Тем не менее ее механизмы пока изучены слабо — в частности, изучение осложнено тем, что существенная часть данных получена на основе моделей травм. Способность полностью дифференцированных клеток к дедифференцировке в нормальных физиологических условиях пока остается невыясненной.
Авторы недавней работы продемонстрировали, что дедифференцировка зрелых соматических клеток может регулироваться митохондриями. В исследовании использовали модельных рыбок данио, в которых определенная популяция эпителиальных клеток помечена генетически. Эпителиальные ионоциты, также известные как NaR-клетки (от Na+-K+-ATPase-rich), экспрессировали GFP. Эти клетки структурно и функционально схожи с клетками кишечного и почечного эпителия человека, в частности, с точки зрения трансклеточного переноса ионов кальция.
При нормальной (0,2 мМ) концентрации Ca2+ в среде, где выращивали зародышей рыб, ионоциты оставались неактивными, однако при переносе в бедную кальцием среду (0,001 мМ) их количество резко увеличивалось. Дальнейший анализ показал, что это происходило именно за счет перехода уже существующих NaR-клеток к активной пролиферации.
Ученые измерили разность потенциалов на мембране митохондрий (ΔΨm), и оказалось, что в делящихся NaR-клетках этот показатель существенно выше, чем в покоящихся. При этом относительное количество мтДНК в клетке не изменялось — это говорит о том, что в пролиферирующих NaR возрастает активность митохондрий, но не митохондриальная масса. Опыты с ингибиторами окислительного фосфорилирования показали, что нарушение функциональности митохондрий препятствует размножению ионоцитов.
Затем авторы изучили, как связаны ΔΨm и сигнальный путь инсулиноподобного фактора роста (IGF-сигналинг). Предыдущие данные указывают на участие сигнального пути IGF-PI3K-Tor в реактивации клеток NaR. Так, их реактивация нарушалась в организме рыбок, дефицитных по гену одного из участников IGF-сигналинга (Papp-aa). Опыты на papp-aa-/- данио показали, что в отсутствие нормального IGF-сигналинга клетки не реагировали на снижение концентрации кальция возрастанием ΔΨm. Напротив, в модели рыбок, дефицитных по кальциевому каналу Trpv6, значение ΔΨm оставалось повышенным даже при физиологическом уровне внеклеточного Ca2+(отсутствие Trpv6 приводит к постоянной активации сигнального пути IGF).
Аналогичные результаты были получены и при обработке зародышей данио ингибиторами ключевых компонентов IGF-сигналинга. Так, воздействие ингибитора рецептора IGF1 или рапамицина препятствовало возрастанию ΔΨm в клетках NaR. Эти данные подтверждают, что активация митохондрий при пролиферации этих ионоцитов регулируется при участии IGF.
Результаты дальнейших экспериментов указывают на роль метаболизма митохондрий в этих процессах. При этом ключевыми для возврата ионоцитов к пролиферации были синтез АТФ и уровень активных форм кислорода (АФК) в митохондриях. Связующим звеном между IGF-сигналингом и АФК оказалась киназа Sgk1. Уровень мРНК sgk1 был намного выше в реактивированных NaR клетках, чем в покоящихся, а нокаут sgk1 при помощи CRISPR/Cas9 нарушал реактивацию клеток.
Авторы работы предположили, что повышенный уровень АФК индуцирует экспрессию или активность Sgk1, что, в свою очередь, влияет на уровень фосфорилирования F1Fo-АТФ-синтазы и стимулирует синтез АТФ. Обработка модельных клеток пероксидом водорода действительно повышала уровень фосфорилирования ATP5B (одной из субъединиц F1-АТФ-синтазы), но этот эффект блокировался ингибитором Sgk1. Полученные данные подтверждают предложенный учеными механизм.
Дополнительно исследователи провели эксперименты на человеческой клеточной линии рака молочной железы MDA-MB-231, руководствуясь предыдущими данными — эта клеточная линия пролиферирует при активации митохондриального метаболизма. Оказалось, что SGK1 в этих клетках также активируется АФК и регулирует клеточный цикл, стимулируя пролиферацию клеток по описанному выше механизму.
Описанный в данной работе механизм связывает сигнальные пути, регулирующие активность митохондрий, с пластичностью эпителиальных клеток. Авторы предполагают, что эта регуляторная система достаточно консервативна, и говорят о необходимости более подробно изучать ее особенности в клетках рака эпителиального происхождения.
Дефекты окислительного фосфорилирования сокращают жизнь клетки
Источник
Li, Y. et al. ROS signaling–induced mitochondrial Sgk1 expression regulates epithelial cell renewal // PNAS (2023). DOI: 10.1073/pnas.2216310120





 0
0