
 Меню
Меню





 Все темы
Все темы
Евгений Кунин: «Почему вирусы архей были изгнаны из эукариотического рая?»
Последний общий предок всех эукариот — LECA — появился примерно два миллиарда лет назад. Сначала бактерия поглотила архею, а затем этот химерный организм поглотил другую бактерию. От первой бактерии эукариоты — и мы с вами — получили мембраны, от археи — ядро, от второй бактерии — митохондрии. Такая схема подтвердилась при анализе вирусов всех трех ветвей эволюции.

Евгений Кунин и его коллеги опубликовали статью «The virome of the last eukaryotic common ancestor and eukaryogenesis» в журнале Nature Microbiology (Mart Krupovic, Valerian V. Dolja & Eugene V. Koonin). В работе проводится анализ виромов различных организмов и делается вывод о происхождении эукариотической клетки путем двух последовательных симбиозов (подробнее на PCR.NEWS). Статья меня ошеломила, поскольку в моих университетских учебниках рисовали совсем другие схемы эукариогенеза. Я попросил Евгения разъяснить возникшие вопросы. Кунин немедленно согласился, предлагаем запись нашей беседы.
Евгений, в познании эукариогенеза я остановился на той стадии, когда было общепринято, что какая-то амебоподобная клетка сожрала бактерию, и из последней получилась митохондрия. А откуда там ядро взялось — ну, как-то само появилось. А когда я стал готовиться к интервью, то понял, что за последние 15 лет появилось много исследований в этом направлении. Ваша работа даже на этом фоне выглядит вызывающе авантюрной. И хотелось бы понять, как ставилась задача, какую гипотезу вы предполагали доказать, и что получилось в результате?
Мы не пытаемся доказать никаких гипотез, это в принципе невозможно. Мы всего лишь проводим некоторое исследование и потом отмечаем, что его результаты существенно лучше совместимы с одним классом гипотез, чем с другими. В нашу исходную задачу вовсе не входило понимание эукариогенеза. Но сложилось так, что из того, что мы делали, произошли, на мой взгляд, довольно существенные выводы, лучше всего совместимые с одним типом гипотез на эту тему.
Тогда расскажите, как формулировалась исходная задача.
Наша с коллегами цель состояла в том, чтобы попытаться реконструировать то, что называется «виром», — разнообразие вирусов, которое было у общего предка эукариот. Это часть довольно большой программы исследований: до этого мы провели аналогичное исследование, пытаясь реконструировать виром общего предка всех клеток, а здесь нам было интересно про эукариот. Важно знать, как жил общий предок всех эукариот, какие у него было вирусы, а их было немало, очевидно. Накапливается много данных относительно геномов разных эукариот, разных вирусов, которые их заражают, о распространении этих вирусов. И поэтому подобные исследования становятся более обоснованными, менее спекулятивными. Этот путь далеко не закончен, но мы прошли уже довольно большое расстояние.
И мы это исследовали самым прямым способом. Вы изображаете эволюционное дерево эукариот, которое довольно хорошо исследовано к нашему времени, и на каждой большой ветке помечаете, какие вирусы заражают данную группу организмов. И далее вы можете проектировать квазиформальными методами — а что было в корне дерева? Очевидно, что если нечто есть почти у всех, в том числе в самых далеких ветвях, отходящих от корня, то, по всей вероятности, это было и в самом начале. Мы провели такое исследование и картировали в этого общего предка всех эукариот довольно большое количество вирусов. Потом мы стали смотреть, каковы возможные предки этих эукариотических вирусов среди вирусов прокариот. И тут мы увидели замечательную вещь, а именно: все эти древние предковые эукариотические вирусы связаны родственными отношениями и, по-видимому, происходят, от вирусов бактерий, а отнюдь не от вирусов архей.
 Евгений Кунин
Евгений Кунин
Вы говорите «все», но, насколько я понял из вашей статьи, кроме одной группы?
Да. В отношении одной важной группы есть проклятая неопределенность, которую тоже легко будет понять в дальнейшем обсуждении. Практически все остальные большие группы предков вирусов эукариот происходят от вирусов бактерий, а не от вирусов архей. Это было интересно и несколько неожиданно осознать. Потому что — что такое вирус? Генетический паразит, который использует информационные системы хозяина, репликацию, транскрипцию, если это ДНК-вирус. Только трансляцию, если это РНК-вирус. Но в любом случае — информационные системы хозяина для репликации собственного генома. Надо сказать, что эти генетические системы у эукариот происходят от соответствующих систем архей, а не бактерий. Это хорошо известно и никем не оспаривается. И поэтому, казалось бы, что и вирусы тоже должны происходить от вирусов архей! Особенно любопытно, что буквально в последние годы были открыты и исследованы непосредственные предки эукариот среди архей, так называемые асгардархеи (подробнее на PCR.NEWS. — Примеч. ред.) Сейчас представляется совершенно определенным, что именно эта ветка архей — непосредственные предки эукариот, точнее, некоторая подветка этой ветки. И удалось в последние годы обнаружить довольно много вирусов, которые заражают эти организмы. Это нетривиально, потому что эти организмы очень плохо растут в культуре, и обнаруживать их вирусы нужно косвенными методами, но на это есть свои приемы. И такие вирусы были исследованы, в одной из таких работ я принимал участие. Многие ждали, и я в том числе, что у этих самых асгардархей будут какие-то интересные вирусы, которые дали начало вирусам эукариот. Оказалось, что ничего подобного! У них вирусы по-своему интересные, но это типичные вирусы архей. А все эукариотические берут свое начало от бактериальных.
И о чем это говорит?
Для начала говорит о том, что эукариоты — это гибридные организмы. Одна часть эукариотических генов происходит от архей, а другая, большая, — от бактерий. Это кажется удивительным, противоречит интуиции, поскольку от архей происходят самые важные, универсальные гены, вся система репликации, транскрипции, трансляции. Но... факты — упрямая вещь, большинство генов эукариот — бактериального происхождения, и мы должны учитывать это в наших попытках реконструировать эволюцию.
Это уже хорошо доказано?
Это не подлежит никакому сомнению. Можно спорить относительно индивидуальных генов, но то, что есть такое разделение, это совершенный факт. А откуда происходят вирусы? По-разному может быть. Может быть, что только от архейных, может быть, что только от бактериальных, может быть, что частично так, частично — эдак. И может быть так, что из других элементов, не вирусных, возникли эти эукариотические виромы. Все четыре возможности находятся «на столе». И так мы это и рассматривали, с открытыми глазами.
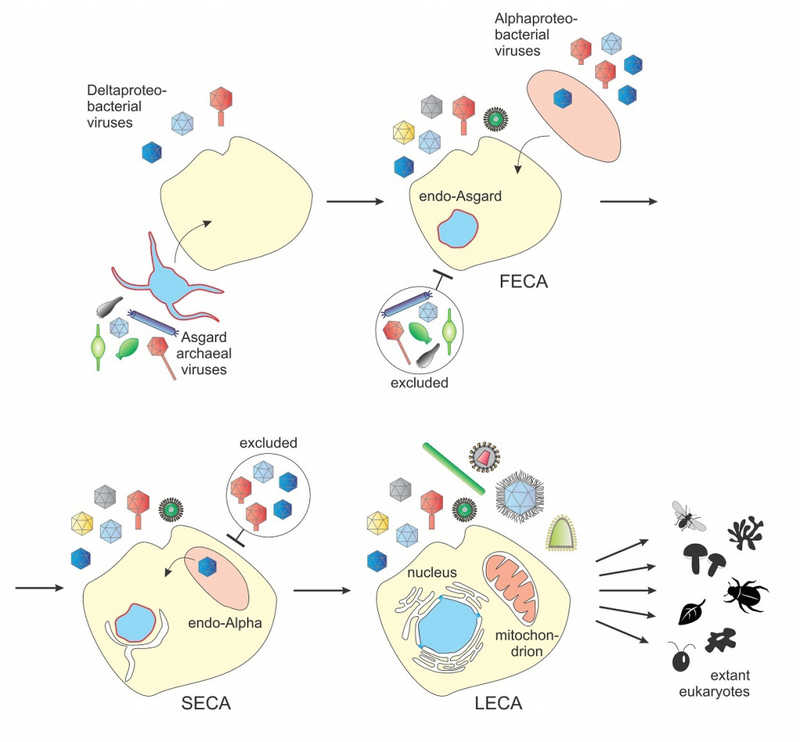
Выясняется, что практически для всех, за одним своеобразным исключением, можно проследить бактериальное происхождение. И мы стали думать, почему это так, почему вирусы архей были изгнаны из эукариотического рая, а бактериальные в него попали? И возникла самая простая мысль, что это связано с устройством эукариотических клеток, а именно — со структурой их мембран. Мембран у эукариот много — цитоплазматическая внешняя мембрана и сложная система внутриклеточных мембран, так называемый эндоплазматический ретикулум, который, в сущности, включают в себя и ядерную мембрану. Все эти мембраны в химическом отношении, в смысле структуры фосфолипидов, из которых они сделаны, являются бактериальными, а не архейными. А там есть принципиальная разница в химических свойствах и в энзимах, которые занимаются их синтезом. Так вот, бактериальные вирусы естественным образом адаптированы миллиардами лет эволюции проникать через бактериальные мембраны, архейные вирусы — через архейные. И мы стали думать, что это обстоятельство и определило структуру вирома эукариот.
Что это значит?
Если сделать еще один шаг, уже теперь в сторону самих стадий эукариогенеза, то возникает идея, что у общего предка всех эукариот была бактериальная мембрана. Как же это могло случиться? Наиболее традиционная гипотеза эукариогенеза включает в себя единственный, очевидный эндосимбиоз: все эукариоты имеют митохондрии, которые являются эндосимбионтами. Самый простой ход мысли состоит в том, что они вторглись в некую предковую клетку. Какую клетку? Довольно долгое время считали, как мы обсуждали в начале разговора, что это был какой-то примитивный одноклеточный эукариот, у него, наверное, уже было ядро, которое своим автогенетическим путем образовалось, и мембраны, и даже способность к фагоцитозу. И вот он захватил эту митохондрию, и все стало уже совсем как теперь. Но потом выяснилось, что никаких эукариот без митохондрий нет, следовательно, симбиоз произошел, по-видимому, на самой ранней стадии.
А тут появились данные о том, что эукариоты в значительной степени происходят от архей, в частности — вся система репликации, транскрипции и трансляции. Потом выяснилось, что не вообще от архей, а от совершенно определенной ветки архей, асгардархей. И тогда появилась модификация гипотезы этого симбиоза, а именно — что такая архея взяла и проглотила альфапротеобактерию, но съесть не сумела, и протеобактерия в ней прижилась в качестве митохондрии. Но тут возникает неловкая ситуация с мембранами: если такое случилось, значит, была снаружи архейная мембрана, внутри — бактериальная, и каким-то образом изнутри эта бактериальная мембрана заместила архейную. Образовала эти внутриклеточные мембраны и заменила всю плазматическую мембрану. Законов физики, которые запрещали бы, чтобы это произошло, не существует, но процесс очень странный.
На самом деле он не более странный, чем тот, который вы описываете в обсуждаемой статье.
Я думаю, что более... И учитывая эту странность, то, что мы поняли про эукариотические виромы, — возникла идея посмотреть на другие гипотезы возникновения эукариот, эукариогенеза. Те, которые постулируют непрерывное существование бактериальных мембран. В первую очередь это так называемый синтрофический сценарий. Согласно этому сценарию, дело происходило наоборот. Бактерия, скорее всего дельтапротеобактерия, заглотила асгардархею. И эта архея стала предком эукариотического ядра.
Эта гипотеза какого года?
Пури Лопеc-Гарсия и Дэвид Морейра впервые опубликовали это в 1998 году. Тогда были не совсем точные представления, основанные на отсутствии данных в то время, но сама идея была предложена именно тогда. А после открытия асгардов и некоторых важных вещей относительно них она была существенно развита и уточнена теми же авторами около трех лет назад. Казалось бы, это более сложный сценарий, но он имеет большие достоинства.
Он же предполагает каким-то образом рассасывание мембраны археи и ее замену на бактериальную мембрану?
Конечно. Ломать — не строить. Это предполагает, что архейная мембрана была потеряна. Она так или иначе была потеряна, что бы ни произошло. Это установленный факт.
 Два симбиоза при происхождении эукариотической клетки. Схема предоставлена Евгением Куниным
Два симбиоза при происхождении эукариотической клетки. Схема предоставлена Евгением Куниным
Почему бы ей не остаться? Ну, были бы мы такие: сверху одно, внутри другое.
Тут мембраны одни, а тут — другие? Однозначного ответа я вам дать не могу. Но это существенное усложнение. Это требует содержать два аппарата для синтеза мембран. Требует, чтобы эти мембраны не смешивались. Физически это возможно, но это не очень хорошо работает. Я думаю, что это было бы крайне неэффективное и неэкономичное решение проблемы. Мы можем об этом порассуждать, но так вышло, что архейная мембрана исчезла. Вопрос не столько «почему?», сколько «как?». И вот разные сценарии эукариогенеза на этот вопрос приводят разные ответы. Надо сказать, что эта гипотеза, которая до открытия асгардархей и иx взаимоотношений с бактериями даже особенно серьезно и не рассматривалась. А тут получила мощную поддержку, которая состоит в том, что эти самые асгардархеи, по крайней мере, многие из них, без бактерий не живут. Образуют то, что называется синтрофические ассоциации с определенными бактериями.
То есть где-то рядом должны находиться бактерии, которые доставляют им определенные вещества?
Не просто «рядом», а в тесных объятиях, в прямом контакте. На электронных микрофотографиях это очень красиво выглядит. Асгарды образуют выросты, которые цепляются не к чему-нибудь, а именно к дельтапротеобактериям. Асгарды производят молекулярный водород, а дельтапротеобактерии его радостно поглощают и восстанавливают сульфат. У этих протеобактерий есть продукты метаболизма, всякая органика, которую получают асгарды в обмен на водород. И тут разговор о таком сценарии эукариогенеза пошел всерьез. Разумеется, этот сценарий подразумевает второй эндосимбиоз, когда химера дельтапротеобактерий и асгарда захватила альфапротеобактерию, которая стала митохондрией.
Здесь консенсус ведь уже давно в науке есть?
В том, что митохондрия есть результат эндосимбиоза, консенсус есть примерно 50 лет. А вот в вопросе о хозяине до сих пор нет. Но сейчас эта синтрофическая гипотеза представляется самой лучшей. Доказательств нет, но свидетельства есть серьезные, может быть, удастся пpонаблюдать что-то подобное экспериментально. Это нелегкие эксперименты, с асгардами очень трудно работать… Может, удастся наблюдать захват, интернационализацию археи дельтапротеобактерией. Пока что мы их наблюдаем в виде эктосимбиоза: они буквально переплетены, но все-таки это две отдельные клетки. То, что мы узнали про вирусы, на мой взгляд, является довольно сильным аргументом с несколько неожиданной стороны в пользу подобных схем происхождения эукариот.
Давайте уточним. Вот эти современные асгарды и современные дельтапротеобактерии — они ведь совсем не те, которые были 4 миллиарда лет назад, я правильно понимаю? Как мы можем экстраполировать?
Во-первых, 4 миллиарда лет назад ничего такого не было вообще. Все эти вещи, эукариогенез, происходили примерно 2 миллиарда лет назад. Теперь, что касается того, что «не такие». Ну, не буквально, но у нас нет основания думать, что они очень сильно изменились. Нет ни малейших оснований думать, что за 2 миллиарда лет существенно изменилась организация геномов и общая организация клеток. И по реконструкции генетического дерева тоже видно, что общие предки всех асгардов были очень похожи на современных. Если бы мы их увидели, то никак не отличили бы.
А зачем вообще такая сложная схема с двойным эндосимбиозом? Почему не предположить, что эукариоты изначально были отдельной ветвью, происходившей прямо от общего предка всех клеточных организмов? Получились бы три ветви: эукариоты, бактерии, археи…
Ну, именно это и предположил в свое время Карл Вёзе, когда открыл архей. Именно он и предложил схему трех доменов. Но жизнь идет и показывает, что это неверно по следующим причинам: потому что есть универсальные гены — универсальные для всей жизни — в первую очередь это гены рибосомного аппарата и всей системы, во вторую очередь там некоторые важные белки, которые принимают участие в транскрипции. И вот если по этим универсальным генам строить, так сказать, дерево жизни, то в этом дереве эукариоты решительно отказываются быть сестринской группой «всех архей». Ну никак! Они оказываются внутри архей. И не просто внутри, а довольно глубоко. Когда асгардархеи были открыты, стало бесспорным, что эукариоты помещаются внутрь асгардов по этим универсальным генам.
Это одно обстоятельство. Теперь второе: я уже упоминал, что помимо генов, унаследованных от архей, у эукариот есть большое количество бактериальных генов. А с ними тоже очень интересно: они разные. Из биологических соображений простейший сценарий мог быть такой: был один эндосимбионт, который дал начало митохондрии, и все бактериальные гены от него и произошли. Так многие думали, и, может быть, думают и теперь. Но с данными это согласуется плохо. Потому что откуда происходят митохондрии, от каких конкретно бактерий, отлично известно, это установил тот же самый Карл Вёзе лет 40 назад. Митохондрии происходят от альфапротеобактерий. Но большинство генов бактериального происхождения, которые есть у эукариот, у альфапротеобактерий никак не прослеживаются.
То есть они не могли быть перенесены из митохондрии в ядро, как раньше рисовали, а пришли еще откуда-то?
Довольно большое количество генов так и было перенесено, да, но не большинство. Знаете, тут есть некоторая неопределенность, которая заключается в том, что у нас нет в руках генома той альфапротеобактерии, которая дала начало митохондриям. Чисто спекулятивно невозможно исключить, что у нее были все эти гены. Но это была бы очень-очень странная альфапротеобактерия. С огромным геномом, в котором встречаются гены, которых у современных альфапротеобактерий нет. И кажется наиболее вероятным, что были и другие бактериальные предки. В частности, есть довольно серьезный сигнал от вышеупомянутых дельтапротеобактерий.
Все же та схема, которая была раньше, радовала тем, что она проста.
Видите ли, как говорится, «у всякой сложной проблемы есть простое, красивое и неверное решение».
Есть же вопросы, на которые не отвечает и сложная схема! Например, куда у эукариот делась последовательность Шайна—Дальгарно? Она есть и у архей, и у бактерий. Другой пример — у эукариот появляется кэп, которого нет ни у архей, ни у бактерий. У архей и бактерий ДНК кольцевая, а у эукариот вдруг появляются линейные хромосомы. Такие примеры можно и дальше перечислять. И, кстати, трехдоменная схема легко бы объясняла это — так изначально было устроено. Можете пару слов сказать?
Много можно сказать. Во-первых, вот что: в реальной жизни все не так, как вы говорите — что у архей и бактерий есть кольцевая ДНК, а у эукариот — нет. Есть немало бактерий, у которых геном состоит из линейной ДНК, а у многих эукариот существуют кольцевые ДНК. Последовательность Шайна—Дальгарно — это небольшое дело. Она короткая и есть далеко не у всех бактерий и архей. Они отлично справляются и без нее. Кэп — да, это важная черта эукариот, но многие бактерии и археи тоже имеют кэпированные мРНК, правда, структура кэпа другая, но он есть. Так что когда начинаешь серьезнее разбираться, то прототипы всего этого уже были. И ничего полностью не исчезло. Но принципиальные решения — да, стали другими. Почему? На этот вопрос однозначных ответов почти никогда нет.
Эволюция так пошла…
Да. Но под действием каких-то факторов, не только случайностей. Возьмем последовательность Шайна—Дальгарно — для чего она нужна? Чтобы обеспечивать трансляцию полицистронных мРНК. Важно то, то последовательность Шайна—Дальгарно находится впереди каждого гена и позволяет рибосомам садиться впереди гена внутри мРНК и начинать оттуда трансляцию. Очень большая доля мРНК у бактерий и архей полицистронные, имеют два-три-четыре гена. У эукариот этого почти нет, подавляющее большинство мРНК кодируют только один ген, и процесс начинается только с определенного конца молекулы мРНК. Почему это так, куда исчезли полицистронные РНК? Исчезли они вместе с оперонами. Полицистронные РНК — это продукт транскрипции оперонов. Почему исчезли у эукариот опероны? Пожалуй, на это вопрос не так сложно ответить: потому что существование оперонов у прокариот в значительной степени поддерживается горизонтальным переносом генов. При переносе генов опероны переносятся как единый кусок. У эукариот после возникновения ядра горизонтальный перенос генов резко снизился. Соответственно, «что с воза упало, то и пропало», — когда мутация разрушает оперон, то воссоздать его уже крайне сложно. Пропадает и потребность иметь полицистронные мРНК. Даже было бы плохо, если бы соседние гены транскрибировались в отдельный транскрипт — тогда совместно регулировалось бы производство совершенно не родственных, никак не взаимодействующих белков. То есть селекция работает против этого.
Да, понятно, почему так получилось. Но есть и другой вопрос. Вы несколько лет назад говорили, что у эукариот большая часть «мусора» остается в геноме. Видно или не видно эти следы? Ту же последовательность?
Последовательность Шайна—Дальгарно — это всего шесть нуклеотидов. Как только они перестают поддерживаться эволюцией, их размывает в мгновение. Мусор остается, но те же силы нейтральной эволюции, которые не дают убрать этот мусор, размывают все следы.
Еще вопрос: есть ведь и другие гипотезы происхождения эукариот, насколько они релевантны тому, что вы видите в генетике?
Те, кто серьезно предлагает какие-то гипотезы, видят в генетике то же, что и мы. Потому что это все опубликовано. Они предлагают это все с учетом тех же данных. Например, есть такая гипотеза «серийного эндосимбиоза», которая постулирует архею как хозяина этого симбиоза, но объясняет разное происхождение бактериальных генов, в частности, довольно позднее происхождение митохондриальных генов серией эндосимбиозов. Хозяева-археи имели довольно хорошую способность захватывать бактерии и делали это несколько раз. И предыдущие эндосимбиозы не прижились как таковые, но оставили после себя гены, а потом — вот, митохондриальный прижился, и это зафиксировало эукариотическую клетку. В этом есть своя разумность, это учитывает данные генетики. Но, на мой взгляд, это хуже согласуется, с одной стороны, с метаболическим симбиозом, а с другой стороны, с эволюцией мембран. И с вирусами.
Вам не удивительно, что столько разных редких событий укладывается в одну такую эволюционную схему?
Что является редким событием? Возникновение эндосимбиоза. А вот возникновение симбиоза вообще, эктосимбиоза — это совсем не редкое явление. Возможностей для попыток возникновения таких химерных организмов сколько угодно. А удачные попытки… Исключать роль случая в эволюции было бы неправильно. Он играл большую роль. Случай подворачивается тем, кто находится в трудной ситуации.
Вам будет интересно




 8075
8075
 0
0










