Меню
Меню





 Все темы
Все темы
Альтернативный путь удлинения теломер в раковой клетке связан с «залипанием» белков на ДНК
Патологическое поддержание длины теломер связано с развитием рака. Авторы статьи в Nucleic Acid Research показали, что альтернативный путь удлинения теломер (ALT), не зависящий от активности теломеразы, включается из-за прочного захвата белков на ДНК и потери функции белка ATRX.
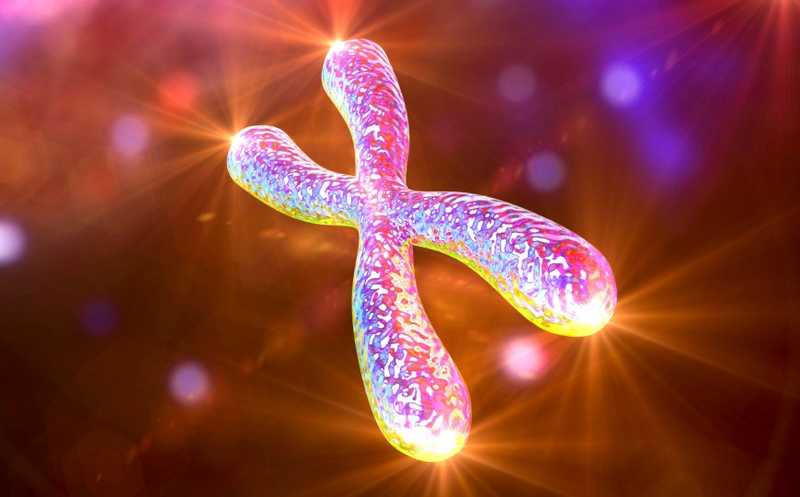
Авторы новой работы, опубликованной в Nucleic Acid Research, показали, что в раковых клетках с дефицитом белка ATRX усиливается связывание белков с ДНК, что приводит к остановке вилки репликации и активирует альтернативный путь удлинения теломер.
При каждой новой репликации эукариотическая ДНК теряет небольшой концевой участок хромосомы, из-за чего число делений клетки ограничено (так называемый лимит Хейфлика). На концах хромосом расположены теломеры — области тандемных повторов TTAGGG общей длиной в тысячи оснований, связанные с белковым комплексом шелтерином.
Огромный интерес к теломерам определяется их участием в процессах старения и развития злокачественных новообразований. Известно, что клетки опухолей способны удлинять свои теломеры. Как правило, это связано с усилением активности теломеразы — специализированного рибонуклеопротеина, который постепенно добавляет теломерные повторы к концам хромосом. Однако при некоторых видах рака клетки используют так называемый альтернативный механизм удлинения теломер (ALT). В его основе лежит аберрантная рекомбинация теломер и консервативный синтез ДНК, известный как репликация, индуцированная разрывом (BIR). Путь ALT характерен для раков мезенхимального происхождения (например, остеосаркоме) и некоторых видов рака центральной нервной системы, таких как глиобластома.
Почти все клетки, у которых отмечен ALT, не способны экспрессировать ген ATRX и связанный с ним DAXX, однако механизмы участия ATRX/DAXX в удлинении теломер пока не ясны.
ATRX — мультифункциональный белок: это фактор ремоделирования хроматина, участвующий в связывании гистонов с определенными участками генома (включая теломерный хроматин), они играет определенные роли в репликации ДНК, а также поддерживает стабильность генома, в том числе стимулируя репарацию двухцепочечных разрывов ДНК.
В новой статье показано, что в клетках с дефицитом ATRX к запуску ALT приводит прочное связывание белков с ДНК. С клеточной ДНК всегда работает множество белков, и эти тесные взаимодействия создают риски образования ковалентных поперечных связей ДНК-белок либо прочного нековалентного захвата белков ДНК. Риски усиливают ультрафиолет, ионизирующая радиация — а также многие химиотерапевтические препараты. (С другой стороны, многие ALT-раки чувствительны к таким препаратам.)
Авторы сравнивали известную линию клеток HeLa с длинными теломерами (в них индуцируется ALT из-за отсутствия гистонового шаперона ASF1) и линии, которые создали они сами с помощью CRISPR/Cas9-нокаута ATRX. Полученные клоны не имели характерных маркеров ALT. Однако под действием пиридостатина (агента, который стабилизирует G4-квадруплексы — вторичные структуры в ДНК) маркеры наблюдались в клетках с нокаутом ATRX, но не в клетках с ATRX дикого типа; также у них появились признаки хрупкости теломер (что может быть связано с застреванием репликативных вилок).
Некоторые другие химиотерапевтические агенты также индуцировали ALT в клетках HeLa, в которых отсутствует ATRX, но не в клонах с ATRX дикого типа. Тот же эффект наблюдался, когда клоны с истощением ATRX обрабатывали формальдегидом, индуцирующим перекрестные связи между белком и ДНК.
С помощью иммунофлюоресцентного анализа FISH авторы подтвердили, что ATRX рекрутируются к богатым GC-парами и повторами областям ДНК (прежде всего теломерам) и участвуют в разрешении неканонических G4-структур в них.
Возникновение неканонических структур ДНК, такие как G4, требуют вмешательства топоизомераз, в частности, топоизомеразы 2А (TOP2А). Ее захват способствует индукции ALT в нокаутированных по ATRX клетках, как и захват ряда других белков, например PARP1 (ключевая молекула ответа на повреждения ДНК). Авторы сочли возможным, что захват топоизормераз и PARP1 на ДНК — решающий этап в инициации ALT.
Развитие ALT также зависит от активности эндонуклеазы MUS81, снижающей избыточную суперспиральность ДНК. Авторы вызвали нокдаун гена, кодирующего MUS81, с помощью малых интерферирующих РНК; клетки с таким нокдауном имели лишь слабые признаки ALT.
Наконец, ученые выяснили, что недостаток ATRX приводит к повышению уровня белков, захваченных на ДНК, и это коррелирует также с активацией ALT. Интересно, что линия с нокдауном ASF1 тоже продемонстрировала увеличение ковалентного захвата TOP1 и одновременную индукцию ALT.
Таким образом, показано большое значение удержания белков на ДНК для альтернативного удлинения теломер при развитии рака. Авторы предлагают модель, согласно которой в отсутствие ATRX или DAXX репликативные вилки становятся нестабильными при встрече с захваченным белком. Таким образом, потеря функции ATRX однозначно опасна, поскольку способствует активации альтернативного пути удлинения теломер.
Источник
Rose, A. M., et al. Induction of the alternative lengthening of telomeres pathway by trapping of proteins onDNA // Nucleic Acids Research (2023). Published online 21 March 2023. DOI: 10.1093/nar/gkad150



 0
0