Меню
Меню





 Все темы
Все темы
«Генетический переключатель» регулирует экспрессию гена Cdx2 в эмбриогенезе
Экспрессия генов в дифференцирующихся клетках эмбриона тонко настраивается, чтобы обеспечить нормальное развитие организма. Ученые из Великобритании выявили молекулярные механизмы, которые регулируют временную экспрессию гена Cdx2 — он необходим для формирования задней части тела у млекопитающих. Авторы показали, как два регуляторных участка в интроне Cdx2 индуцируют активацию и затем выключение этого гена, чтобы обеспечить правильное развитие осевого скелета.
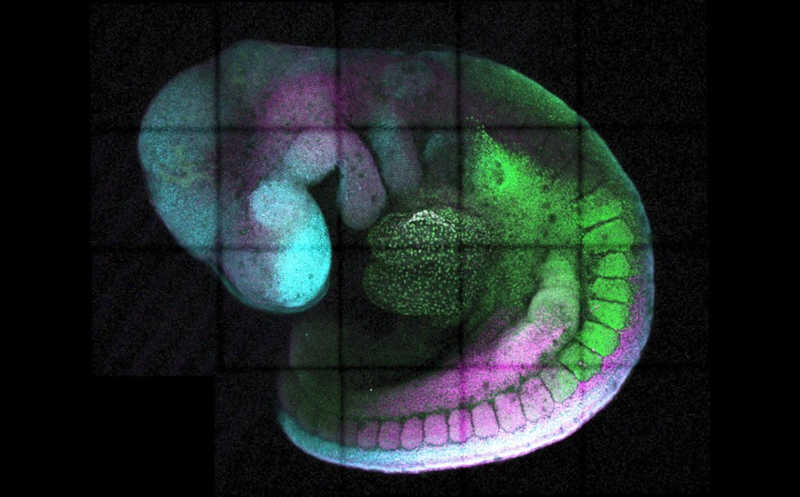
Разнообразие клеточных типов в организме млекопитающих объясняется специфичными профилями экспрессии генов, обуславливающими клеточную идентичность, — это результат активности генных регуляторных сетей. Особенно важна регуляция экспрессии на этапе дифференцировки клеток, в частности, при эмбриональном развитии, когда клетки-предшественники дают начало всем типам тканей организма. В формировании нервной системы важен гомеобоксный ген Cdx2 — при его участии из клеток каудального эпибласта (CEpi) в течение нескольких дней формируется будущий спинной мозг, после чего ген Cdx2 «выключается». Авторы публикации в Developmental Cell выяснили, как контролируется своевременное ограничение экспрессии Cdx2 — они обнаружили цис-регуляторный элемент, который за это отвечает.
Исследователи выявили три цис-регуляторных элемента (промотор Cdx2, интронный элемент и 5′-участок), которые доступны в CEpi-подобных клетках (CEpiL-клетки), экспрессирующих Cdx2, но теряют доступность в предшественниках спинного мозга. Цис-регуляторные элементы, активные в других тканях, не демонстрировали такой динамики экспрессии. Анализ доступности хроматина проводили методом ATAC-seq в дифференцирующихся стволовых клетках in vitro и на эмбриональных клетках in vivo.
Чтобы определить роль отдельных CRE в регуляции Cdx2 при формировании каудальной части тела, авторы получили линии эмбриональных стволовых клеток с нокаутом того или иного регуляторого элемента. Экспрессия Cdx2 резко снижалась при удалении интронного элемента или промотора, что указывает на ключевую роль интронного участка в инициации экспрессии Cdx2. При этом регуляторная активность интронного CRE оказалась тканеспецифичной и не обнаруживалась в нейромезодермальных клетках-предшественниках. Делеция 5′ CRE не влияла на начальную экспрессию Cdx2 в CEpiL, но приводила к ее снижению на поздних стадиях. Так авторы показали, что различные CRE по-разному регулируют запуск и поддержание Cdx2 в зависимости от клеточного контекста.
Проанализировав интронный элемент, ученые обнаружили в нем два участка (P1 и P2), которые выполняют противоположные функции. В CEpiL-клетках активный участок P1 связывается с коактиватором p300 и фактором LEF1, способствуя индукции Cdx2, тогда как в плюрипотентном состоянии клетки участок P2 связывается с репрессорами CTNNB1, TCF3 и HDAC3. Удаление P1 подавляло индукцию Cdx2 аналогично удалению всего интрона, тогда как удаление P2 не мешало активации, но приводило к продлению экспрессии Cdx2 в предшественниках спинного мозга. Таким образом, P1 необходим для активации, а P2 — для временного ограничения экспрессии Cdx2.
Дальнейший анализ показал, что P1 и P2 содержат различные сайты связывания ретиноевой кислоты (RA): P1 содержит мотивы RXRG (Retinoid X Receptor Gamma), активные в CEpiL-клетках, тогда как P2 содержит репрессорный мотив RARB (Retinoic Acid Receptor Beta), активирующийся при дифференцировке в клетки спинного мозга. Удаление или замена RARB на RXRG в P2 усиливали и продлевали экспрессию Cdx2, указывая на ключевую роль состава RA-распознающих мотивов. Предполагается, что RARB опосредует выключение Cdx2 через взаимодействие с факторами LEF1 и SOX2, которые менее активно связывались с P2 при удалении из него этого мотива.
Чтобы подтвердить роль элемента P2 in vivo, авторы с помощью CRISPR-Cas9 вырезали RARB-мотив из этого участка в эмбрионах мыши. Полученные фенотипы — удлиненный передний конец спинного мозга, аномальная структура хвостовой почки — соответствовали последствиям избыточной экспрессии Cdx2. Таким образом, P2 и его RARB-мотив необходимы для своевременного подавления Cdx2 и правильного формирования задней части тела.
Опубликованная работа подчеркивает, как важен точный контроль экспрессии для дифференцировки клеток. Она показывает, как экспрессия Cdx2 своевременно усиливается и «выключается» при участии тандема энхансерного участка P1 и аттенюатора P2, соответственно. Результаты могут помочь и в идентификации многих других цис-регуляторных элементов: «...предыдущие исследования показывают, что наш геном может содержать множество различных типов элементов, которые тонко настраивают экспрессию генов, но их нелегко идентифицировать. Если мы сможем решить эту проблему, это откроет огромный потенциал для открытия новых способов лечения заболеваний путем тонкой настройки экспрессии генов там и тогда, где и когда это необходимо», – комментирует доктор Вики Метзис, руководитель исследования.
Транспозоны играют роль альтернативных промоторов во время раннего эмбриогенеза млекопитающих
Источник
Irene Amblard, et al. A dual enhancer-attenuator element ensures transient Cdx2 expression during mouse posterior body formation // Developmental Cell. Published online 27 June 2025. DOI: 10.1016/j.devcel.2025.06.006
Цитата по пресс-релизу
Вам будет интересно

 125
125
 0
0


 157
0
157
0