Меню
Меню





 Все темы
Все темы
Барьерный иммунитет легких регулируют антигенпрезентирующие эпителиальные клетки
Эпителиальные клетки легких экспрессируют главный комплекс гистосовместимости 2 (MHC-II). MHC-II — это белковый комплекс, который презентует антигены клеткам иммунной системы. Эпителиальные клетки способствуют активации тканевых T-клеток памяти в ответ на повторную инфекцию. Отсутствие экспрессии MHC-II в эпителиальных клетках легких мыши нарушает работу барьерного иммунитета.
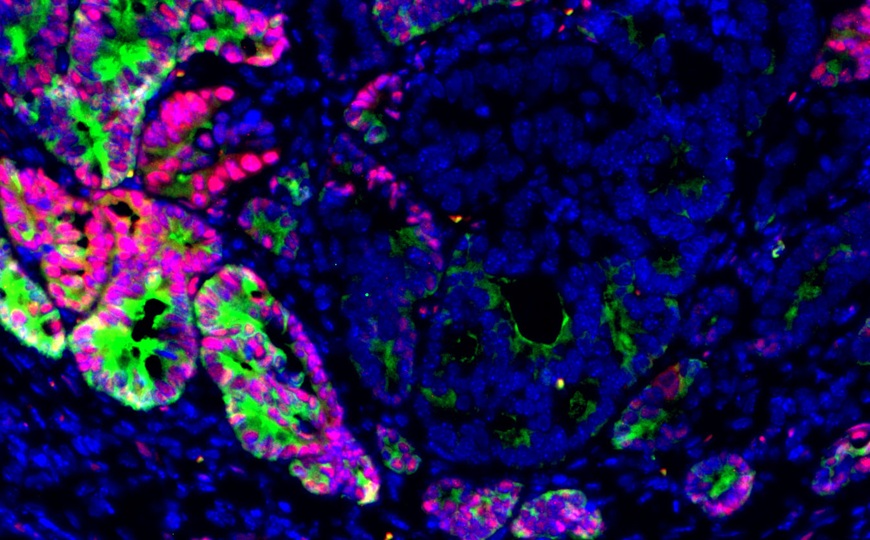
Тканевые Т-клетки памяти (Trm) — одна из первых линий обороны против инфекции в пограничных тканях, таких как кожа и легкие. Корректная работа Trm позволяет организму быстро и эффективно реагировать на различные антигены, которые неизбежно входят в контакт с барьерными тканями. Группа ученых из США показала, что презентация антигенов эпителиальными клетками легких играет ключевую роль в работе барьерного иммунитета.
Первым делом ученые исследовали экспрессию главного комплекса гистосовместимости 2 (MHC-II) в эпителиальных клетках легких (LEC). MHC-II — это белковый комплекс, который обеспечивает презентацию антигенов иммунным клеткам. Анализ показал, что экспрессия MHC-II в LEC повышается после инфицирования мышей Streptococcus pneumoniae. Ученые также вывели линию мышей, у которых MHC-II в LEC не экспрессировался (MHC-IIΔEpi). У таких мышей была нарушена активация CD4+ Trm в легких в ответ на повторную инфекцию, что подтверждает предположение о том, что LEC действуют как антигенпрезентирующие клетки.
Презентацию антигенов модулируют другие сигналы, костимуляторы, поэтому авторы изучили экспрессию других генов в LEC. В зависимости от паттернов экспрессии эпителиальные клетки легких разделили на кластеры. После заражения с течением времени часть кластеров появлялась и исчезала. Экспрессия также зависела от типов клеток. Так, клетки проводящих воздушных путей в определенных условиях экспрессировали MHC-II и постоянно — костимуляторы. LEC альвеол постоянно экспрессировали MHC-II и PD-L1, после инфекции уровень их экспрессии возрастал.
Авторы предположили, что анатомическое разделение LEC с различными паттернами презентации антигенов ведет к формированию «ниш» CD4+ Trm, локализованных в определенных частях легких. Для проверки этой гипотезы они отслеживали динамику CD4+ Т-лимфоцитов легких после многократных заражений в тех временных промежутках, когда LEC презентуют антигены. Через 14–24 дня после заражения, когда организм подавил инфекцию, число CD4+ Т-клеток сокращалось в паренхиме легких, но не вдоль воздухоносных путей. Вокруг этих путей обнаруживались «ниши» Т-клеток с постоянными популяциями. Через 35 дней после заражения все лимфоциты в «нишах» представляли из себя CD69+ Trm.
Результаты указывают на то, что клеточное окружение вокруг воздухоносных путей способствует формированию CD4+ Trm. Чтобы показать, как LEC влияют на этот процесс, ученые провели повторный опыт, на этот раз сравнивая мышей MHC-IIΔEpi и животных дикого типа. У мышей MHC-IIΔEpi было меньше очагов агрегации Т-клеток в эпителии воздухоносных путей. При этом динамика численности лимфоцитов в паренхиме не менялась.
Далее ученые исследовали фенотипическое разнообразие CD4+ Trm-клеток. Они проанализировали экспрессию транскрипционных факторов, определяющих направление развития Т-клеток (lineage-defining transcription factors, LDTF). CD4+ T-клетки разделили на кластеры по профилям экспрессии. Из 20 клестеров 17 экспрессировали по крайней мере два LDTF. Изменения, происходящие с кластерами у MHC-IIΔEpi мышей, демонстрируют, что MHC-II в LEC ограничивает пластичность и мультипотентность CD4+ Тrm-клеток. Ученые также показали, что отсутствие MHC-II влияет на соотношение различных Т-клеток при иммунном ответе.
Авторы сравнили динамику численности различных иммунных клеток в легких у MHC-IIΔEpi мышей и контролей. Спустя восемь часов после заражения MHC-IIΔEpi мышей наблюдали повышенное число эозинофилов и макрофагов, но сниженную численность моноцитов и произошедших от них иммунных клеток. Подобные изменения схожи с теми, что происходят у пациентов, проходящих лечение ингибиторами контрольных точек иммунного ответа. Обратив на это внимание, ученые проанализировали экспрессию PD-L1 и обнаружили, что его уровень коррелирует с уровнем MHC-II в LEC. Также альвеолярные LEC у MHC-IIΔEpi мышей не экспрессируют PD-L1. То есть MHC-II в LEC направляет адаптивный иммунный ответ как через презентацию антигенов, так и через PD-L1.
Таким образом, новая работа наглядно демонстрирует важную роль, которую играют эпителиальные клетки легких в иммунном ответе. Ученые считают, что дальнейшее изучение презентации антигенов этими клетками может способствовать более глубокому пониманию иммунологических процессов, ассоциированных с хроническими заболеваниями, раком и терапии иммуномодуляторами.
Источник
Shenoy A.T., et al. Antigen presentation by lung epithelial cells directs CD4+ TRM cell function and regulates barrier immunity // Nature Communications 12, 5834 (2021), published: October 05, 2021, DOI: 10.1038/s41467-021-26045-w




 0
0