Меню
Меню





 Все темы
Все темы
Дефицит MDGA2 у мышей приводит к появлению симптомов расстройства аутистического спектра
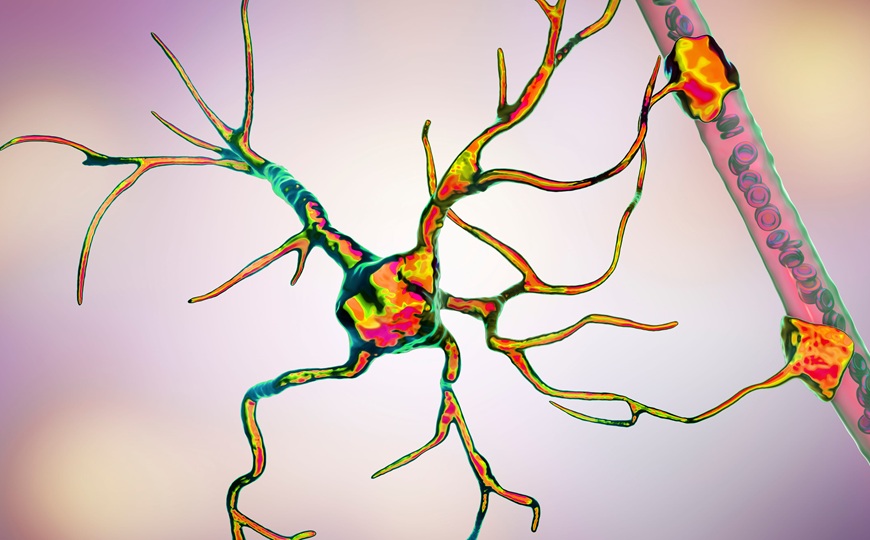
Исследователи из Китая обнаружили, что симптомы расстройств аутистического спектра (РАС) у мышей связаны с дисбалансом между белками MDGA2 и BDNF, которые играют важную роль в формировании и работе синапсов. Дефицит белка MDGA2 приводил к гиперактивации сигнального пути BDNF/TrkB, из-за чего увеличивалась как частота, так и амплитуда возбуждающих сигналов в гиппокампе, появлялись признаки РАС. Подавление активности рецептора TrkB различными способами снижало частоту стереотипного поведения и улучшало социальные взаимодействия у мышей с нокаутом MDGA2.

Расстройства аутистического спектра (РАС) — это нарушения нейроразвития, затрагивающие примерно 1% населения Земли. Несмотря на все усилия, этиология РАС все еще не до конца изучена. Было выявлено более 100 генов, ассоциированных с этими состояниями, включая гены SHANK, DLG4, NRXN и NLGN, кодирующие синаптические белки. Недавно у пациентов с РАС были найдены мутации с потерей функции и миссенс-мутации в гене MDGA2, однако его функции при этих состояниях неизвестны.
В исследовании, опубликованном в PLOS Biology, ученые из Китая проанализировали роль белка MDGA2 в регуляции нейрональной активности, а также его связь с развитием РАС. MDGA2 — это мембранный белок, находящийся в области синаптических контактов и регулирующий синаптическую передачу. В отличие от своего гомолога MDGA1, который преимущественно модулирует ГАМКергические синапсы через нейролигин 2, MDGA2 взаимодействует с нейролигином 1 и рецептором TrkB, контролируя глутаматергическую передачу. Оба гена возникли в результате дупликации у позвоночных, но MDGA1 более консервативен и экспрессируется в организме шире, чем MDGA2, который обнаруживается только в мозге. Для MDGA1 была найдена ассоциация с шизофренией, когда как о MDGA2 было известно довольно мало.
TrkB является основным рецептором BDNF (нейротрофического фактора мозга), конкурентным ингибитором которого оказался белок MDGA2. До этой работы уже была известна связь BDNF с РАС: у пациентов были обнаружены изменения концентрации BDNF в сыворотке крови, а также однонуклеотидные полиморфизмы в его гене. Однако данные были противоречивыми: хотя большинство работ указывало на усиление BDNF/TrkB-сигналинга при РАС, некоторые исследования на животных моделях показывали, что активация этого пути, наоборот, смягчает аутистические симптомы.
В новой работе авторы использовали генетически модифицированных мышей с нокаутом гена MDGA2. Поведенческие тесты показали, что такие животные демонстрируют признаки, схожие с РАС у людей: сниженный социальный интерес в трехкамерном социальном тесте (животные проводили значительно меньше времени с другой мышью, предпочитая оставаться в одиночестве), стереотипное поведение (повторяющийся груминг) и повышенную тревожность в тесте с открытым полем (животные избегали открытых участков и демонстрировали характерное замирание при перемещении в новые зоны).
Кроме того, в мозге этих мышей выявили изменения синаптической активности в гиппокампе и префронтальной коре, областях, связанных с социальным поведением и когнитивными функциями. В этих зонах отмечалась повышенная активность возбуждающих синапсов, связанная с усилением сигнального пути BDNF/TrkB. Авторы проанализировали возбуждающие и тормозные постсинаптические сигналы и обнаружили, что у нокаутных мышей значительно увеличилась как частота, так и амплитуда возбуждающих сигналов.
Ученые предположили, что MDGA2 и BDNF конкурируют за связывание с TrkB, а сдвиг этого баланса приводит к нарушению нейроразвития. Исследователи разработали синтетический пептид MDGA2-P2, имитирующий участок MAM-домена белка MDGA2. Этот пептид вводили в мозг мышей с нокаутом MDGA2, что значительно повлияло на их состояние: у животных снизилась частота стереотипного поведения, улучшились социальные взаимодействия, а электрофизиологические исследования показали нормализацию синаптической активности в префронтальной коре и гиппокампе. Механизм действия пептида заключался в селективном блокировании взаимодействия BDNF с рецептором TrkB, что предотвращало его чрезмерную активацию и восстанавливало баланс возбуждающих и тормозных синаптических сигналов. При этом пептид не влиял на общую двигательную активность.
Кроме того, ученые исследовали и ингибитор TrkB — соединение ANA-12. Хотя ANA-12 снижал фосфорилирование TrkB, его эффект на поведение был не таким значительным. Как и пептид MDGA2-P2, ANA-12 улучшал социальные взаимодействия, но не влиял на повторяющееся поведение. В то время как ANA-12 обеспечивал быстрое, но временное подавление гиперактивности TrkB, пептид MDGA2-P2 обладал более устойчивым эффектом, вероятно, за счет восстановления естественного механизма регуляции через MDGA2.
Таким образом, авторы раскрыли новый механизм, ассоциированный с РАС, связывающий дефицит белка MDGA2 с гиперактивацией BDNF/TrkB-сигнального пути и последующими нарушениями синаптической пластичности.
Нокаут единственного гена вызывает у мышей признаки расстройства аутистического спектра
Источник:
Zhao D., et al. (2025) Mdga2 deficiency leads to an aberrant activation of BDNF/TrkB signaling that underlies autism-relevant synaptic and behavioral changes in mice. // PLoS Biol 23(3): e3003047. DOI: 10.1371/journal.pbio.3003047
Вам будет интересно

 41
41
 0
0


 36
0
36
0