Меню
Меню





 Все темы
Все темы
Диатомовые водоросли заняли новую экологическую нишу благодаря бактериальному гену
Большинство диатомовых водорослей — фотосинтетики, они производят до 40% атмосферного кислорода. Однако некоторые представители рода Nitzschia извлекают энергию из полисахаридов, произведенных многоклеточными водорослями, то есть являются гетеротрофами. Международный коллектив ученых показал, что водоросли получили гены ферментов, необходимых для гетеротрофного питания, от морских бактерий путем горизонтального переноса и последующих мутаций.
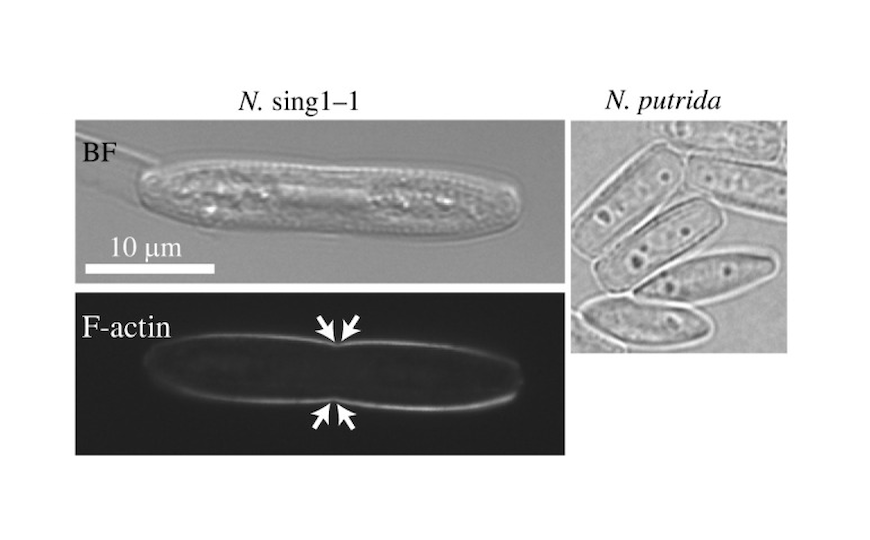
Нефотосинтезирующие диатомовые водоросли из Сингапура.
Credit:
Open Biol. 2023. DOI: 10.1098/rsob.230148 | CC BY 4.0
Симбиоз фотосинтезирующей цианобактерии с эукариотическим гетеротрофом около 1,5 млрд лет назад стал началом эволюции эукариот, использующих энергию света для синтеза органики, — фотоавтотрофов. Первое эндосимбиотическое событие привело к появлению наземных растений, зеленых и красных водорослей и глаукофитов (их хлоропласты похожи на цианобактерии). В эволюционной истории водорослей эндосимбиоз был не единственным: некоторые их таксоны имеют хлоропласты с тремя или четырьмя мембранами, которые, вероятно, происходят от поглощенных зеленых или красных водорослей. Однако и обратный переход, с с автотрофного типа питания на гетеротрофный, происходил среди самых разных групп водорослей.
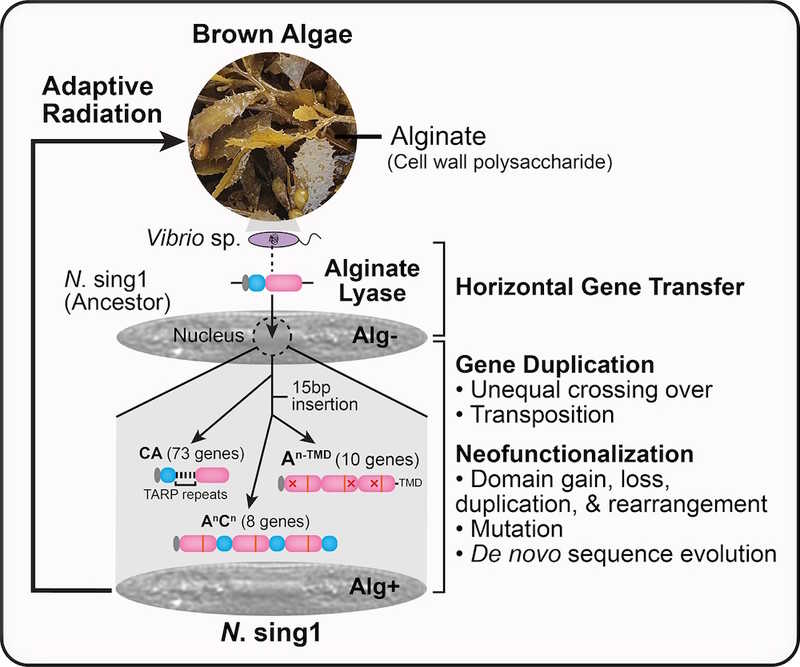
Гетеротрофные диатомовые водоросли, принадлежащие к роду Nitzschia, обитают на поверхности многоклеточных водорослей или в разлагающемся растительном материале. Исследователи из Сингапура с коллабораторами из Германии и Норвегии выяснили, как группа диатомовых водорослей перешла с фотоавтотрофного питания на гетеротрофное и заняла новую экологическую нишу.
Ученые выполнили полногеномное секвенирование изолята нефотосинтетической диатомовой водоросли Nitzschia sing1 с сингапурского острова Сентоса. Они собрали и аннотировали геном и провели в нем поиск генов углевод-активных ферментов (CAZyme), которые позволяют усваивать полисахариды других водорослей. Эти гены сравнили с профилями CAZyme других диатомовых водорослей, как фотосинтетических, так и гетеротрофных. Исследователи обратили внимание, что N. sing1 кодирует 91 ген, содержащий хотя бы один домен альгинатлиазы (ALY) из семейства полисахаридлиаз 7 — это необычно много. (Альгинаты — полисахариды клеточной стенки красных, бурых и зеленых водорослей.) При этом 77 из 91 генов помимо каталитического домена ALY (А-домен) кодировали домены углевод-связывающего модуля 32 (С-домен), обеспечивающие специфичность фермента к конкретному углеводороду.
Анализ последовательностей этих генов показал, что их можно разделить на три семейства: гены, кодирующие концевой С-домен и А-домен (семейство СА), гены, кодирующие исключительно тандемные повторы А-доменов (семейство An-TMD), и гены, кодирующие комбинации A- и С-доменов, также в виде тандемов (семейство AnCn).
Филогенетический анализ выделенных генов семейства СА выявил монофилетический характер этой группы. Это значит, что все их разнообразие возникло в ходе эволюции из одного гена. Чтобы выяснить, откуда появился предковый ген, ученые сравнили последовательности белков ALY N. sing 1 c белками других организмов и выяснили, что их «ближайший родственник» — род морских бактерий Vibrio. Эти данные указывают на то, что каталитический домен ALY был приобретен N. sing1 в результате горизонтального переноса генов от морской бактерии. Гены семейства CA похожи на «предка» больше всего.
Далее авторы реконструировали эволюцию An-TMD и AnCn. Оба семейства произошли от СА в результате вставок в А-домен, дупликаций и потерь доменов и появления тандемных повторов.
Морские бактерии утилизируют полисахариды, выделяя эндолитические (расщепляющие внутренние связи в цепочке) альгинатлиазы и затем транспортируя олигосахаридные фрагменты внутрь клетки, где экзолитические олигоальгинатлиазы отщепляют от них концевые моносахара. Но у N. sing 1 нет гомологов транспортеров или олигоальгинатлиаз, следовательно, они усваивают полисахариды с помощью иных механизмов.
Каталитическую активность белков с ALY-доменами изучали in vitro — оценивали разжижение альгинатного геля экстрактами Escherichia coli, экспрессировавших тот или иной белок. Чтобы понять, как влияют на ферментативную активность мутации, приобретенные N. sing 1, авторы провели анализ пространственной структуры белков при помощи AlphaFold2. Результаты показали, что последовательность альгинатлиазы, специфичная для семейства AnCn обладает экзолитической активностью (достаточно низкой), в то время как СА сохранил предковый эндолитический механизм действия.
 Credit: Jedd Group |
Credit: Jedd Group |
Как было упомянуто, ферменты семейства An-TMD не имеют углевод-связывающего домена. А в последовательности каталитического домена у них присутствуют мутации, дезактивирующие его. Поэтому ученые предположили, что белок этого семейства заякоривается в мембране и служит рецептором для внутриклеточного транспорта альгинатов. Флуоресцентный анализ указал на смену локализации белка от клеточной мембраны к лизосомам, что подтверждает данное предположение. Вероятно, N. sing 1 поглощает продукты внеклеточного расщепления альгинатов из водорослевой биомассы, и дальнейшая их обработка происходит путем эндоцитоза, внутри вакуоли.
Семейство CA содержит специфические TARP-повторы, не содержащиеся в бактериальных ALY. Эти повторы богаты положительно заряженной аминокислотой аргинином и способствуют связыванию отрицательно заряженных альгинатов, что также было подтверждено флуоресцентной микроскопией.
Таким образом, некоторые диатомовые водоросли перешли к гетеротрофному типу питания благодаря горизонтальному переносу генов, а последующая диверсификация функций предкового гена помогла им освоить новую экологическую нишу.
Эволюция со скоростью света: как ИИ помог «разогнать» фотосинтез
Источник
Zeng Hao Lim, et al. Diatom heterotrophy on brown algal polysaccharides emerged through horizontal gene transfer, gene duplication, and neofunctionalization // PLoS Biology 23(4), published online 1 April 2025. DOI: 10.1371/journal.pbio.3003038




 0
0