Меню
Меню





 Все темы
Все темы
Гликированный CD43 защищает раковые клетки от врожденного и приобретенного иммунитета
Наличие белка CD47 на поверхности опухолевых клеток — это сигнал «не ешь меня», который подавляет противоопухолевую активность макрофагов. Но нейтрализующие антитела к CD47 продемонстрировали плохие результаты в КИ. Исследователи из США и Польши показали, что CD47 препятствует фагоцитозу мышиными макрофагами, но не человеческими. А для человеческих макрофагов может стать препятствием другой белок, CD43. После гликозилирования и сиалирования он формирует гликоиммунный барьер, который защищает раковые клетки не только от макрофагов, но и от T- и NK-клеток.
Макрофаги фагоцитируют или не трогают клетки, подчиняясь сигналам «съешь меня» и «не ешь меня» на их поверхности. Белок CD47 на поверхности опухолевых клеток — важный сигнал «не ешь меня», который препятствует противоопухолевой активности макрофагов. Однако в фазе 3 клинических испытаний нейтрализующие антитела к CD47 для лечения миелодиспластических синдромов и острого миелоидного лейкоза (ОМЛ) показали ограниченную клиническую эффективность. Исследователи из США и Польши выяснили, какие еще сигналы раковых клеток могут препятствовать фагоцитозу.
Клетки ОМЛ (MV411 и MOLM13) кокультивировали с макрофагами в течение 18–24 часа. После этого клетки разделили и подсчитали. Фагоцитоз без стимула проходил не очень активно, но под действием IFNγ процесс усиливался в 4–6 раз. CRISPR-скрининг провели с помощью Cas9 и Cas12a. Через 18 часов кокультивирования количество клеток и гидРНК анализировали.
Как и ожидалось, делеция гена APMAP уменьшала фракцию клеток ОМЛ и обогащала фракцию макрофагов. Еще были выявлены гены, вовлеченные в O-связанное гликозилирование (SPN, C1GALT1 и C1GALT1C1), TNFα-сигналинг (PPP6C) и эндосомальный транспорт (VPS25), а также специфичные для клеточной линии регуляторы, включая CDC7, ATP6V1A, MZT1 и VPS29 в MV411 и FADD, IKBKG, MAP3K7, и NHLRC2 в MOLM13.
Делеция CD47 не делала фагоцитоз более активным. Аналогично, делеция QPCTL, фермента, модифицирующего CD47 и необходимого для оптимального связывания CD47 с SIRPα, не усилила фагоцитоз. В то же время в опытах на мышиных клетках делеция CD47 активировала фагоцитоз.
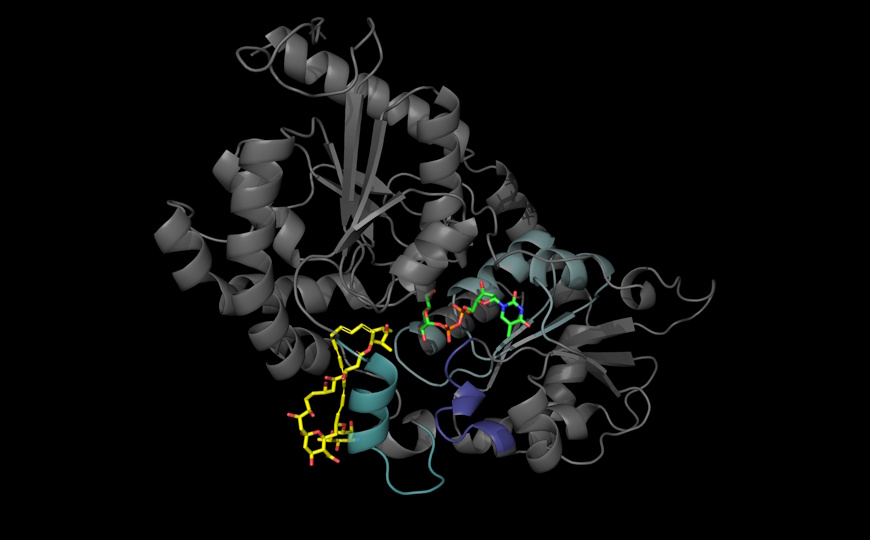
В целом, PPP6C, SPN, APMAP, VPS29 и NHLRC2 были основными отрицательными регуляторами антителонезависимого фагоцитоза, а NLRC5, TAPBP, TAP2, PPP6C и PTPN6 — антителозависимого. Отрицательными регуляторами обоих процессов были APMAP, TMEM30A, гены пути O-связанного гликозилирования (C1GALT1, C1GALT1C1, SLC39A9, SLC35A2) и SPN, кодирующий CD43, гликопротеин клеточной поверхности, подвергающийся обширному O-гликозилированию.
O-связанное гликозилирование — посттрансляционная модификация клеточной поверхности и секретируемых белков, которая присоединяет гликозидные цепи, заканчивающиеся сиаловой кислотой. C1GALT1 кодирует Т-синтазу, которая катализирует ранний этап O-гликозилирования. C1GALT1C1 является белком-шапероном, необходимым для стабильности C1GALT1. SLC39A9 регулирует экспрессию C1GALT1, а SLC35A2 транспортирует уридиндифосфат (УДФ)–галактозу, которая включается в структуры O-гликанов.
При нокауте C1GALT1 и C1GALT1C1 раковые клетки росли с той же скоростью, что и клетки дикого типа, но были больше подвержены антителозависимому и антителонезависимому фагоцитозу. При этом факторы, выделяемые макрофагами, не были ответственны за повышение чувствительности к фагоцитозу. Делеция SLC39A9 или SLC35A2 также активировала фагоцитоз. Те же результаты были получены в опытах на мышах.
Авторы предположили, что эффект от делеции C1GALT1/C1 может быль опосредован потерей конечного остатка сиаловой кислоты. Действительно, обработка клеток ферментом сиалидазой от Vibrio cholerae тоже повышала их чувствительность к фагоцитозу.
O-гликозилирование идет по остаткам серина и треонина, так что авторы оценили содержание этих аминокислотных остатков в поверхностных белках. CD43 (SPN) получил высший балл, так как его внеклеточный домен более чем на 30% состоит из серина и треонина. Экспрессия CD43 на раковых клетках подавляла фагоцитоз in vitro и in vivo. Дальнейшие опыты подтвердили, что O-связанное гликозилирование ингибирует фагоцитоз раковых клеток макрофагами посредством посттрансляционного добавления ингибирующих концевых остатков сиаловой кислоты к CD43.
CD43 оверэкспрессируется в клетках лейкемии человека, что не характерно для здоровых гемопоэтических клеток. Анти-CD43 антитела усиливали фагоцитоз раковых клеток макрофагами. Примечательно, что CD43 защищал клетки лейкемии не только от макрофагов, но и от T- и NK-клеток.
Таким образом, гликопротеин CD43 является мощным ингибитором врожденного и адаптивного противоракового иммунитета. Ингибирующая активность CD43 в отношении иммунных клеток зависит от посттрансляционных модификаций, которые добавляются посредством O-связанного гликозилирования и сиалирования. Сиалированный CD43 является потенциальной терапевтической мишенью для лечения ОМЛ.
Блокировка опухолевого сигнала «не ешь меня» повысила эффективность иммунотерапии рака
Источник:
Jooho Chung, et al. Sialylated CD43 forms a glyco-immune barrier that restrains antileukemic immunity // Science (2026), published 09 April 2026, DOI: 10.1126/science.ady5196

Вам будет интересно

 93
93
 0
0


 92
0
92
0
 112
0
112
0
 131
0
131
0