Меню
Меню





 Все темы
Все темы
Гранзим К запускает четвертый путь активации системы комплемента при ревматоидном артрите
Группа ученых из США открыла новый потенциальный путь активации системы комплемента — через гранзим К. Эта протеаза выделяется в основном эффекторными Т-клетками и, как выяснилось, может активировать весь каскад комплемента. Особую роль она играет при воспалительных заболеваниях: например, при ревматоидном артрите уровень гранзима К в синовиальной оболочке суставов возрастает и активирует систему комплемента, приводя к системному воспалению.
Система комплемента — это часть врожденного иммунитета, состоящая из примерно 50 белков плазмы крови. Существует три пути ее активации — классический, альтернативный и лектиновый, — которые запускают каскад протеолитических реакций. В итоге происходят выработка анафилотоксинов C3a и C5a — они способствуют хемотаксису иммунных клеток, — опсонизацияклеток патогенов и образование мембраноатакующиего комплекса (МАК), который встраивается в мембраны бактериальных клеток и разрушает их. Группа ученых из США открыла четвертый возможный путь активации системы комплемента — через гранзим К, который вырабатывается преимущественно Т-клетками. Этот путь активации комплемента вносит значительный вклад в воспалительную реакцию при ревматоидном артрите и других заболеваниях, связанных с системным воспалением.
Гранзимы — это семейство сериновых протеаз, которые используются для уничтожения опухолевых или зараженных вирусом клеток. Однако функции гранзима К, активность которого похожа на триптазу (протеаза–маркер активации тучных клеток), до сих пор оставались неясными. Уже установлено, что гранзим К в больших количествах экспрессируется CD8+ Т-клетками, находящимися в воспаленных тканях.
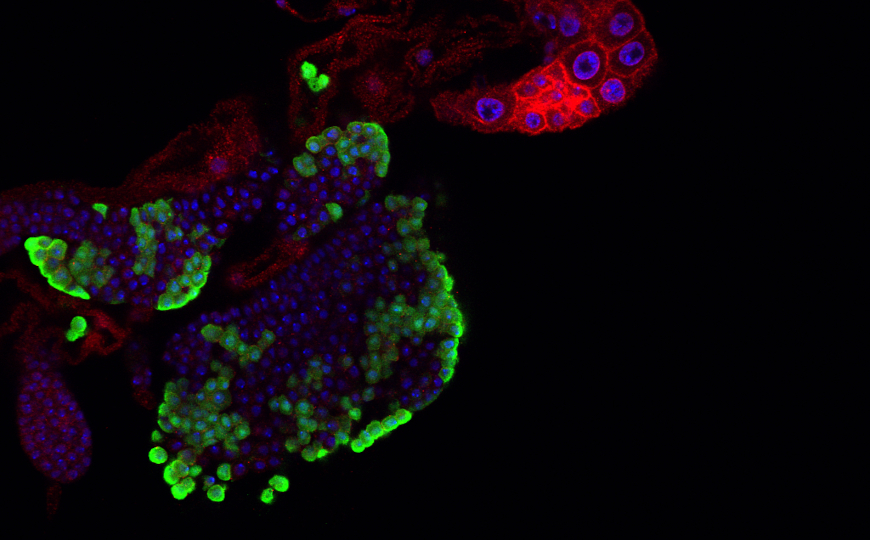
Чтобы подробнее охарактеризовать экспрессию гранзима К, ученые проанализировали наборы данных секвенирования РНК. Они показали, что в крови гранзим К продуцируется преимущественно CD8+ Т-лимфоцитами, а также MAIIT-клетками (инвариантные Т-клетки, связанные со слизистыми оболочками), NK-клетками, некоторыми CD4+ Т-клетками. В синовиальной оболочке суставов пациентов, больных ревматоидным артритом или остеоартритом, гранзим К также в основном секретировался CD8+ Т-клетками, в меньшем количестве — CD4+ Т-клетками, NK-клетками и γδ Т-клетками. Те же клетки экспрессировали гранзим К и в других тканях пациентов с хроническими воспалительными заболеваниями: ревматоидным артритом, волчанкой, язвенным колитом, болезнью Крона.
Более детальный анализ показал, что CD8+ Т-клетки выделяют гранзим К постоянно. Даже в отсутствие стимуляции Т-клеточного рецептора (TCR) эти клетки выделяли гранзим К. При этом стимуляция TCR незначительно влияла на его экспрессию, тогда как другие гранзимы (например, гранзим B) начинают секретироваться только при стимуляции TCR.
Из-за этого факта ученые предположили, что субстрат гранзима К постоянно находится во внеклеточном пространстве. Чтобы определить, как именно действует гранзим К, авторы идентифицировали его гомологи — оказалось, что он обладает высокой гомологией не только с другими гранзимами, но и с протеазами, запускающими каскад системы комплемента: с фактором D, C1s и MASP-1. Из-за этого авторы статьи задались вопросом, может ли гранзим К тоже активировать систему комплемента.
В альтернативном пути активации системы комплемента происходит спонтанный гидролиз молекулы C3, и одна из получившихся частей — C3b — связывается с фактором B, который разрезается фактором D. В итоге получается фрагмент Bb и образуется C3-конвертаза альтернативного пути — C3bBb. Ученые инкубировали гранзим К с фактором B и C3b, но, в отличие от фактора D, не наблюдали образования Bb.
В классическом и лектиновом путях активации системы комплемента C1s или MASP-1 разрезает C2 и C4, в результате образуется C3-конвертаза — C2aC4b. Когда гранзим К инкубировали с C4, он, как и C1s, разрезал его до C4b. В присутствии C4 гранзим К также мог разрезать C2 с образованием C2a. Эти фрагменты, в свою очередь, объединялись в С3-конвертазу, способную разрезать C3, в то время как сам по себе гранзим К не был способен действовать на C3.
Анализ данных секвенирования РНК показал, что фибробласты способны секретировать компоненты системы комплемента C2, C3 и C4. В синовиальной ткани больных ревматоидным артритом именно они были основными источниками этих компонентов. При этом выработка этих белков повышалась под действием интерферона-гамма и TNF, секретируемых CD8+ Т-клетками.
Активность C3-конвертазы, разрезающей C3, ученые подтвердили по активности образуемого C3a — он способен воздействовать на тучные клетки, приводя к их дегрануляции. Эта дегрануляция происходила при инкубации тучных клеток с гранзимом К, а также C2, C3 и C4.
Белки C4b и C3b также называются опсонинами: они содержат активную тиоэфирную связь, за счет которой «облепляют» поверхность клеток патогенов и привлекают к ним фагоциты. Если C4b или C3b находятся в растворе, а не вблизи поверхности, тиоэфирная связь быстро инактивируется водой, и белки обезвреживаются, чтобы они не взаимодействовали с клетками хозяина. Соответственно, для эффективной опсонизации протеазы, активирующие систему комплемента, также должны быть прикреплены к мембранам клеток. С использованием проточной цитометрии ученые подтвердили, что гранзим К может связываться с мембранами клеток эндотелия, фибробластов и моноцитов, активируя систему комплемента и способствуя опсонизации. Более подробный анализ показал, что гранзим К связывается с мембранами за счет электростатического взаимодействия с отрицательно заряженными сульфатированными гликозаминогликанами, т.е. для взаимодействия гранзиму К не требуется специфических рецепторов.
C3b, который образуется под действием C3-конвертазы, входит в ее состав и образует C5-конвертазу. Она расщепляет C5 с образованием C5a, еще одного анафилотоксина, и C5b, который затем связывается с C6, C7, C8, C9 и образует МАК. При инкубации гранзима К с клетками эндотелия человека и C2, C3, C4, C5 действительно образовывалась С5-конвертаза. Вырабатывался C5a, который успешно активировал C5aR1-рецептор на клетках. При добавлении в реакцию также C6, C7, C8, C9 образовывался МАК, что подтверждали проточной цитометрией. Таким образом, эксперименты in vitro доказали, что гранзим К способен запускать полный каскад системы комплемента.
Далее исследователи показали вклад системы комплемента, активируемой гранзимом К, в воспаление при ревматоидном артрите. В образцах синовиальной оболочки пациентов гранзим К колокализовался с C3b и C5a, что указывает на причину воспаления в этих тканях.
Роль гранзима К дополнительно доказали in vivo, используя две мышиные модели воспалительных заболеваний человека — ревматоидного артрита (модель AIA) и псориаза (модель дерматита, вызванного имихимодом). При нокауте Gzmk — гена, кодирующего гранзим К — у мышей с ревматоидным артритом значительно снизилась отечность суставов, а также упал уровень C3d и C4d — стабильных продуктов распада C3b и C4b. У мышей со смоделированным псориазом при нокауте Gzmk также снизилась краснота кожи и шелушение, уменьшился уровень белков системы комплемента. Таким образом, гранзим К может стать мишенью для терапии, направленной на снижение патологического воспаления.
Два гена, связанные с апоптозом, участвуют в патогенезе ревматоидного артрита и остеопороза
Источник

Вам будет интересно
 116
116
 0
0