Меню
Меню





 Все темы
Все темы
Митомейоз — способ получения половых клеток из соматических
В Nature Communications опубликована статья о получении оплодотворяемых яйцеклеток из фибробластов кожи человека. Это подтверждает принципиальную возможность применения такого подхода при некоторых случаях бесплодия. Ранее исследователи получили живых мышей из гаплоидных яйцеклеток, содержащих генетический материал соматических клеток и оплодотворенных сперматозоидами in vitro. Однако для адаптации этой технологии к человеку необходимы дополнительные исследования.
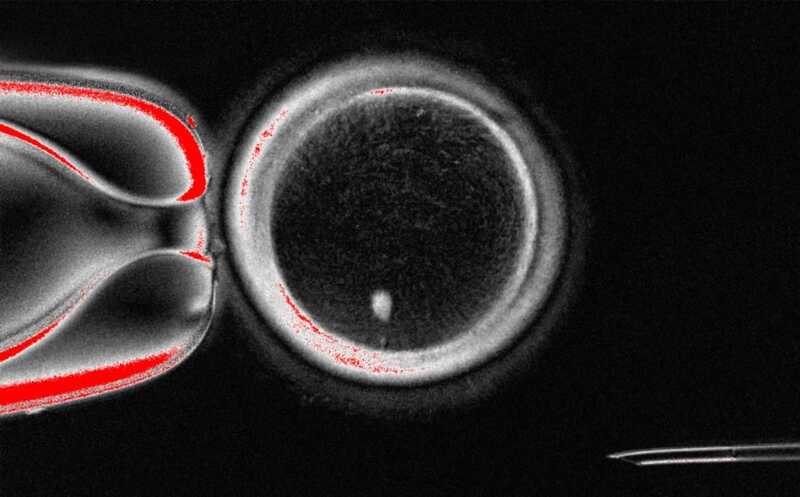
Человеческий SCNT-ооцит с веретеном деления (белая точка)
Credit:
Mitalipov laboratory | Пресс-релиз
Ученые из Орегонского университета здравоохранения и науки под руководством Шухрата Миталипова применили к человеческим клеткам необычную технологию, которая позволяет получить из диплоидной соматической клетки гаплоидную яйцеклетку.
От бесплодия страдают миллионы людей в мире, и многие ее случаи связаны с отсутствием у пациента функциональных половых клеток (яйцеклеток или сперматозоидов). Так, у женщин старше 35 лет снижение количества и качества ооцитов становится одной из ведущих причин бесплодия. При этом снижается и вероятность успешного исхода экстракорпорального оплодотворения (ЭКО).
Методы получения половых клеток из соматических клеток (in vitro гаметогенез) активно исследуются. У мышей возможно получить ооциты и сперматозоиды из плюрипотентных половых клеток. Однако оогенез у человека — сложный и длительный процесс, который начинается еще во внутриутробном периоде; ооцит завершает свое развитие более чем за десять лет, к началу половой зрелости. Подобные сроки неприемлемы для клинического применения.
Функциональные ооциты могут также быть получены путем переноса генома соматической клетки в донорский ооцит (somatic cell nuclear transfer, SCNT). При этом собственные цитоплазматические факторы ооцита перепрограммируют соматический геном, поддерживают индукцию тотипотентности и развитие эмбриона после оплодотворения.
Перед овуляцией диплоидный первичный ооцит вступает в мейоз, который задерживается в метафазе II (MII) и завершается, когда происходит оплодотворение. Донорские ооциты, таким образом, находятся в MII, их цитоплазма сохраняет метафазную активность. Изначально применялись подходы, при которых SCNT-ооцит переходил к эмбриональному развитию без оплодотворения и завершения мейоза. Таким способом были получены клонированные млекопитающие, а также эмбриональные стволовые клетки человека (ЭСК) из соматических.
Однако для лечения бесплодия нужны гаплоидные ооциты, подобные естественным, которые могут быть оплодотворены сперматозоидом. Исследователи из Орегонского университета здравоохранения и науки вместе с коллегами из Южной Кореи ранее провели уменьшение плоидности в мышином SCNT-ооците и после его оплодотворения и имплантации самкам-реципиентам получили живых мышей, которые сами дали потомство.
В новой работе Шухрат Миталипов с коллегами адаптировали эту процедуру для SCNT-ооцитов человека. Ооциты с диплоидным геномом соматической клетки проходили деление, которое авторы назвали «митомейоз», так как оно сочетает в себе черты митоза и мейоза.
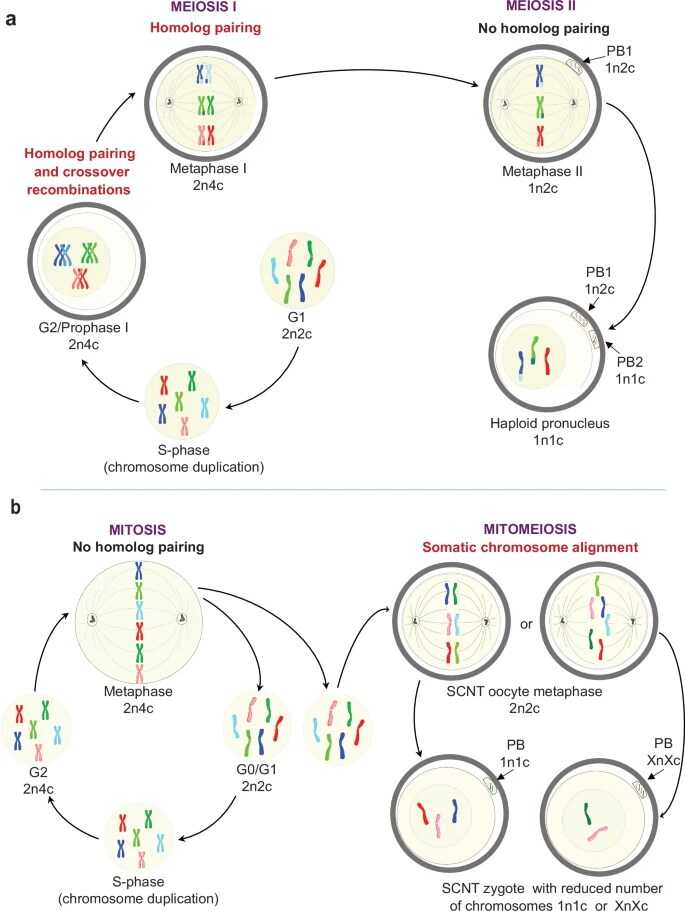 Естественный мейоз, митоз и митомейоз. PB — полярные тельца, в которых направляются «лишние» хромосомы в ходе оогенеза, чтобы получить гаплоидный пронуклеус ооцита; *n — число наборов хромосом, *c — число копий генома в клетке. Как и мейоз, митомейоз приводит к редукции плоидности (2n à 1n). Однако к полюсам веретена деления расходятся не сестринские хроматиды, а гомологичные хромосомы соматической клетки. При этом возможны сбои и появление клеток с аномальным количеством хромосом. Credit: Nat Commun (2025). DOI:
10.1038/s41467-025-63454-7 |
CC BY-NC-ND 4.0
Естественный мейоз, митоз и митомейоз. PB — полярные тельца, в которых направляются «лишние» хромосомы в ходе оогенеза, чтобы получить гаплоидный пронуклеус ооцита; *n — число наборов хромосом, *c — число копий генома в клетке. Как и мейоз, митомейоз приводит к редукции плоидности (2n à 1n). Однако к полюсам веретена деления расходятся не сестринские хроматиды, а гомологичные хромосомы соматической клетки. При этом возможны сбои и появление клеток с аномальным количеством хромосом. Credit: Nat Commun (2025). DOI:
10.1038/s41467-025-63454-7 |
CC BY-NC-ND 4.0
В метафазе митоза и в метафазе II мейоза к полюсам клетки расходятся сестринские хроматиды, возникшие при репликации ДНК, — идентичные при митозе, идентичные за исключением участков кроссинговера при мейозе. Однако в метафазе SCNT-ооцита положение вдоль экватора занимают пары гомологичных хромосом, полученных индивидом от отца и от матери. Ранее авторы показали, что в клетках инбредных (генетически близких) мышей при этом довольно часто происходит правильная сегрегация хромосом — после деления образуются полные гаплоидные геномы.
Для исследования на человеческих клетках использовали ооциты здоровых доноров-добровольцев после обычной процедуры, применяемой в клинике. Из них удаляли веретено деления, затем проводили слияние энуклеированного ооцита с фибробластом кожи в фазе клеточного цикла G0/G1. При дальнейшем культивировании клеток в течение 1-2 часов в них появлялись веретена деления, как с нормальной, так и с аномальной морфологией.
Контрольные ооциты и SCNT-ооциты оплодотворяли сперматозоидами in vitro с помощью интрацитоплазматической инъекции (ИКСИ). Около 80% контрольных оплодотворенных ооцитов выделили полярное тельце и образовали женский и мужской пронуклеусы, однако лишь около четверти SCNT-ооцитов выделили полярное тельце, еще меньшее количество сформировало пронуклеусы, и эти процессы протекали существенно медленнее. Четверть их прошла деление, но остановилась на стадии двух клеток. Следовательно, естественной стимуляции входа в метафазу при оплодотворении SCNT-ооцита может быть недостаточно.
Остановка мейоза в ооцитах поддерживается высокой активностью фактора, способствующего созреванию (MPF), который включает в себя циклинзависимую протеинкиназу 1 (Cdk1) и циклин B. При оплодотворении ооцита изменение концентрации ионов кальция приводит к разрушению циклина B и инактивации Cdk1. Исследователи протестировали различные режимы инактивации MPF и подобрали оптимальный, который включал повышенный уровень Ca2+ в среде, электропорацию и инкубацию с росковитином, ингибитором Cdk1.
Когда 82 оплодотворенных SCNT-ооцита обработали таким образом, большая их часть стала выделять полярные тельца и проходить дальнейшие деления. Около 9% к шестому дню культивирования достигли стадии бластоцисты.
Авторы проверили, как происходит сегрегация соматических хромосом в SCNT-ооцитах при митомейозе, в дополнительном изящном эксперименте. К исследованию привлекли донора фибробластов — женщину, родители которой различались по этническому происхождению (что позволяло надежно отличать ее гомологичные хромосомы), и неродственного донора спермы. Это давало возможность определять происхождение гомологичных хромосом в зиготе с помощью секвенирования. Таким образом проанализировали 90 эмбрионов. Почти у половины все хромосомы оказались в пронуклеусе или в полярном тельце, у остальных паттерны сегрегации варьировали от 3:43 до 23:23. В среднем около 10 хромосом (максимум 16) разделились правильно, то есть одна из двух гомологичных хромосом попала в пронуклеус, другая в полярное тельце. Такая картина соответствует случайному распределению.
Тем не менее SCNT-зиготы могли проходить последующие митотические деления. В некоторых хромосомы SCNT-ооцита и сперматозоида интегрировались в единый геном, и все бластомеры демонстрировали один и тот же хромосомный набор, другие зиготы оказались мозаичными, с различным составом хромосом в бластомерах.
Каким образом можно повысить при митомейозе выход гаплоидных яйцеклеток с точной сегрегацией хромосом, мы спросили у руководителя работы Шухрата Миталипова, директора Центра клеточной и генной терапии эмбрионов Орегонского университета здравоохранения и науки.
— Мы уже пару лет работаем над этим, изучаем, как это происходит при природном мейозе. Главное требование, чтобы гомологичные хромосомы нашли друг друга и спарились по всей длине в профазе митомейоза. При природном мейозе это, видимо, достигается индукцией десятков запрограммированных двунитевых разрывов (double strand breaks, DBS) в одной из каждых гомологичных хромосом. Это привлекает комплексы репарации с участием гомологичной рекомбинации, которые и ищут, находят и спаривают гомологичные хромосомы. DSBs репарируются, при этом происходят кроссоверы, а дальше уже спаренные хромосомы входят в метафазу и потом расходятся после оплодотворения формируя галоидный геном. Мы будем модифицировать митомейоз и следовать по тому же механизму. Все очень просто.
Шухрат Миталипов: «Нужна эффективность редактирования не менее 90%»
Источник
Marti Gutierrez, N., et al. Induction of experimental cell division to generate cells with reduced chromosome ploidy // Nature Communications. 16, 8340 (2025). DOI: 10.1038/s41467-025-63454-7

Вам будет интересно

 62
62
 0
0


 52
0
52
0