Меню
Меню





 Все темы
Все темы
Обратный ход РНК-полимеразы регулирует экспрессию генов в клетках млекопитающих
РНК-полимераза II способна проскальзывать назад по матричной цепи, высвобождая 3’-конец недосинтезированной РНК — это явление называется обратным ходом. Коллектив ученых из США картировал такие события в клетках млекопитающих. Исследователи показали, что обратный ход случается не только в участках, где полимераза не может проехать дальше — он также контролирует экспрессию многих ключевых белков, участвующих в регуляции клеточного цикла и развитии тканей. Например, в клетках, где ход полимеразы возобновиться не может, сильно снижается экспрессия гистонов.
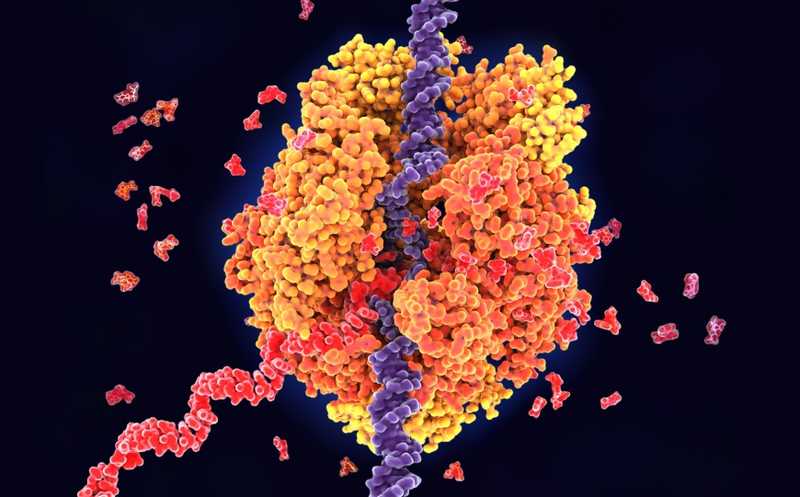
РНК-полимераза II производит все транскрипты белок-кодирующих генов и многие некодирующие РНК у эукариот. В 1997 году Евгений Нудлер и его коллеги опубликовали работу , в которой показали, что РНК-полимераза может иногда проскальзывать назад по матричной цепи — это явление они назвали «обратным ходом» или «обратным смещением» (англ. backtracking). Позднее это явление связали с регуляцией генов и репарацией ДНК. Теперь же Нудлер и соавторы подробно картировали события обратного хода в клетках млекопитающих и описали их регуляторную роль.
События обратного хода могут разрешаться при участии фактора транскрипции TFIIS. Однако это происходит не во всех случаях — вариант, когда РНК-полимераза останавливается на более продолжительное время, прежде чем продолжить транскрипцию, назван персистентным обратным ходом.
Для картирования сайтов персистентного обратного хода исследователи разработали метод LORAX-seq (long-range cleavage sequencing). Он позволяет напрямую детектировать фрагменты РНК, стабильно получающиеся при обратном ходе РНК-полимеразы, по всему геному человека. Чтобы выявить такие РНК, из ядер клеток выделяют хроматин, после чего обрабатывают их ДНКазой. Транскрипционные комплексы иммунопреципитировали и очищали от 5′-фосфатов. Затем их инкубировали с TFIIS in vitro для расщепления и элюирования фрагментов РНК, полученных при обратном ходе полимеразы. Из целевых фрагментов готовили библиотеки для секвенирования.
После картирования ученые обнаружили, что полученные обратным ходом РНК в основном имеют длину от 20 до 40 нт. 3′-конец этих последовательностей был расположен вблизи точек остановки РНК-полимеразы, вычисленных по данным mNET-seq (секвенирование нативных удлиняющихся транскриптов млекопитающих). Однако не все выявленные в анализе остановки полимеразы приводили к ее обратному ходу. Участки, для которых было характерно это явление, располагались преимущественно в промоторах генов и вблизи сайтов сплайсинга.
Авторы статьи предположили, что РНК-полимераза II может обнаруживаться в позициях, расположенных вблизи идентифицированных точек остановки. Чтобы это проверить, они адаптировали метод секвенирования нативных удлиняющихся транскриптов, добавив в него этап обработки РНКазой I, а также обеспечив захват транскрипционных комплексов при помощи 4-тиоуридина — это необходимо, чтобы предотвратить перемещение РНК-полимеразы в ходе пробоподготовки. Ожидалось, что в этом эксперименте РНКаза I переварит высвободившийся при обратном ходе конец РНК, что приведет к смещению детектируемых следов сдвига.
Авторы сравнили сигнал пауз, приводящих к обратному ходу полимеразы (сигнал LORAX-seq) с общими данными сильных пауз (mNET-seq). Анализ показал, что 3′-концы фрагментов РНК, детектированные вышеописанным методом, лежали вблизи участков сильных пауз, тогда как в участках обратного хода сигнал 3′-концов РНК смещался на 20 нт. Такое изменение наблюдалось в масштабах всего генома. Исследователи, однако, отмечают, что более короткие фрагменты РНК, полученные обратным ходом РНК-полимеразы, вероятно, также присутствуют в клетках, но не детектируются секвенированием.
Затем исследователи перешли к анализу функциональной роли персистентного обратного хода РНК-полимеразы в различных генах. Они сравнили клетки дикого типа с клетками, нокаутными по TFIIS. Оказалось, что гены с наиболее сильным сигналом LORAX-seq, то есть те гены, для которых места обратного хода РНК-полимеразы были наиболее характерны, экспрессировались значительно слабее в клетках с нокаутом этого фактора транскрипции. Таким образом, обратный ход полимеразы снижал экспрессию генов в тех случаях, когда он не разрешался при участии этого транскрипционного фактора.
Анализ генной онтологии выявил, что гены, содержащие участки персистентного обратного хода РНК-полимеразы, связаны с трансляцией, сборкой нуклеосом и регуляцией клеточного цикла. Также среди них встречались гены, кодирующие белки теплового шока и некоторые ключевые факторы транскрипции, такие как ATF4 и HOXB9.
Чтобы изучить влияние неразрешенного обратного хода на гистоны, ученые синхронизировали клеточный цикл в культуре, а затем измерили экспрессию мРНК гистона H2AC20 методом количественной ПЦР с обратной транскрипцией (RT-qPCR). В клетках дикого типа экспрессия этого гистона возрастала в пять раз через четыре часа после выхода из синхронизации, тогда как в клетках, нокаутных по TFIIS, — менее чем в два раза. Таким образом, события обратного хода накапливаются на H2AC20, и для правильного уровня экспрессии во время деления они должны разрешаться при участии TFIIS.
Полученные данные подтвердились и методом секвенирования РНК, которым авторы воспользовались, чтобы проверить, будет ли такой эффект наблюдаться на геномном уровне. В клетках дикого типа экспрессия генов гистонов значительно возрастала через четыре часа после выхода из синхронизации. При этом в нокаутных клетках за это время не повышалась экспрессия ни одного гистона.
Разработанный Нудлером и соавторами метод LORAX-seq позволил с высокой точностью картировать участки в геноме млекопитающих, где происходят события персистентного обратного хода РНК-полимеразы. Анализ выявил регуляторную роль этих событий в экспрессии ключевых генов, участвующих в регуляции клеточного цикла и деления, а также в развитии тканей. Это, в частости, может объяснять, почему летальна делеция фактора транскрипции TFIIS, необходимого для разрешения остановок РНК-полимеразы.
Источник
Kevin B. Yang, et al. Persistence of backtracking by human RNA polymerase II. // Molecular Cell, 2024. DOI: 10.1016/j.molcel.2024.01.019

Вам будет интересно

 130
130
 0
0


 158
0
158
0