Меню
Меню





 Все темы
Все темы
Почему разорванные фрагменты хромосомы остаются вместе
Хромотрипсис — это патологическая фрагментация хромосом, после которой фрагменты снова соединяются, но уже с ошибками, что может способствовать малигнизации клетки. В новой работе описан белковый комплекс, который удерживает фрагменты рядом друг с другом.

Авторы новой статьи, опубликованной в Nature, исследовали механизмы, благодаря которым удерживаются вместе фрагменты хромосом, возникающие во время хромотрипсиса.
Хромотрипсис — явление, характерное для большинства опухолей, но описанное лишь в 2011 году: одномоментная фрагментация хромосом и последующая их неточная сборка. Это катастрофическое разрушение происходит в течение одного клеточного деления. Одна или даже несколько хромосом разрываются десятки или даже сотни частей, которые не деградируют далее, а снова собираются, но уже в измененном порядке и с потерей участков. Хромотрипсис зачастую стимулирует развитие рака — из-за потери онкосупрессоров, амплификации онкогенов (в том числе при участии внехромосомных кольцевых ДНК) и онкогенных слияний хромосом.
Хромотриптические хромосомы происходят из неправильно сегрегированных хромосом или хромосомных фрагментов, инкапсулированных в микроядрах — аномальных внеядерных структурах. Условия в микроядрах способствуют фрагментации: ДНК в них хуже защищена, чем в ядре, отсутствуют системы репарации.
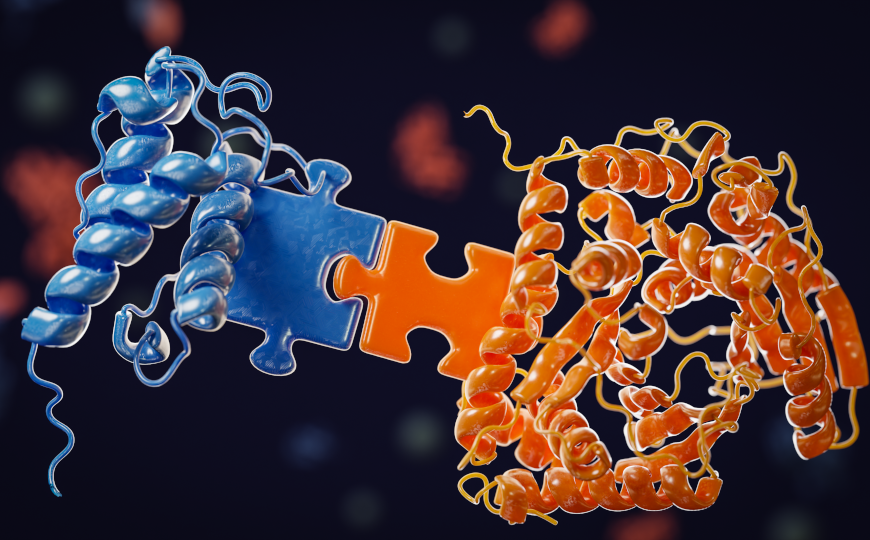
Существовало представление о том, что фрагменты хромосом, не содержащие центромер, распределяются по дочерним клеткам случайным образом. Однако теперь установлено, что они преимущественно попадают в одну дочернюю клетку. Авторы нового исследования впервые описали молекулярные механизмы, благодаря которым это происходит. Фрагменты разорванных хромосом удерживает вместе комплекс белков, включающий в себя медиатор чекпоинта повреждения ДНК 1 (mediator of DNA damage checkpoint 1, MDC1), связывающий топоизомеразу II белок (topoisomerase II-binding protein 1, TOPBP1) и клеточный ингибитор PP2A (CIP2A).
Ученые фрагментировали хромосомы в клетках RPE, нокаутированных по гену онкосупрессора p53, а также клетки DLD1 с мутациями в этом гене. В других экспериментах дефицит p53 также вызывали с помощью малых интерферирующих РНК.
Хромотрипсис и образование микроядер (микронуклеацию) вызывали с помощью ингибитора протеинкиназы Mps1, а для формирования митотических хроматиновых мостиков и катенированных (сцепленных) сестринских хроматид клетки обрабатывали ингибитором топоизомеразы II. Поскольку находящиеся в микроядрах хромосомы распадаются перед митозом или сразу после вхождения митоз, деление клеток останавливали с помощью ингибитора кинезина Eg5. Фрагменты разорванных хромосом отслеживали благодаря характерному фосфорилированию гистона H2AX по положению S139A (γH2AX), которое возникает неподалеку от двухцепочечных разрывов ДНК.
В клетках DLD1 сегрегация Y-хромосомы нарушалась за счет инактивации ее центромеры. В следующем митозе появлялись Y-хромосомы, положительные по γH2AX. Во всех вариантах эксперимента фрагменты хромосом оставались рядом, часто даже после множественных митозов.
Авторы предположили, что фрагменты могут удерживаться вместе за счет взаимодействия с PolD3, субъединицей полимеразы дельта (ранее было показано, что в поврежденных хромосомах во время митоза идет активный синтез ДНК). Действительно, PolD3 участвовала в репарации поврежденных хромосом, но на попадание фрагментов в одни и те же микроядра ее присутствие или отсутствие не влияло.
Выяснилось, что белки MDC1, TOPBP1 и CIP2A активно рекрутируются к областям, содержащим γH2AX. Эксперименты с РНК-интерференцией подтвердили, что снижение концентрации любого из этих трех белков приводит к случайному распределению частей хромосом. Эффект был связан не с увеличением числа разрывов, а с потерей комплексов, удерживающих эти фрагменты вместе. MDC1 взаимодействует с участками двухцепочечных разрывов и в свою очередь рекрутирует TOPBP1 и CIP2A.
Наконец, полногеномное секвенирование 24 клонов клеток с микронуклеацией выявило существенное увеличение в них числа структурных вариантов генома и копийности повторов ДНК. Исследование экспрессии генов в более чем 8 тысячах геномов реальных опухолей показало, что в них усилена экспрессия CIP2A и TOPBP1, а активность кодирующих их генов обратно коррелирует с числом копий повторов.
CIP2A не является необходимым для деления клеток, его экспрессия низка в большинстве нормальных тканей человека. Авторы считает, что это привлекательная мишень для терапии хромосомно нестабильных опухолей. Нарушение кластеризации приводит к попаданию фрагментов хромосом в цитоплазму, что запускает защитный ответ с участием сигнальных путей врожденного иммунитета и снижает жизнеспособность раковой клетки.
Недавно в Nature была опубликована статья другой научной группы, которая также подтвердила роль комплекса CIP2A-TOPBP1 в удержании вместе фрагментов хромосом, возникших при хромотрипсисе. Авторы этой работы оценили вклад MDC1 как незначительный.
Источник
Trivedi, P., et al. Mitotic tethering enables inheritance of shattered micronuclear chromosomes // Nature (2023). Published: 14 June 2023. DOI: 10.1038/s41586-023-06216-z



 0
0