
 Меню
Меню





 Все темы
Все темы
Алексей Шацких: «Транспозоны — это мусорная ДНК или склад полезных функций?»
Очередная лекция блока «Геном» научно-просветительского проекта «Метафаза» от PCR.NEWS и парка «Зарядье» посвящена мобильным генетическим элементам. Они не только вредны, но и полезны для организмов: «прирученные» транспозоны могут принимать участие в реализации ряда функций, таких как формирование долговременной памяти, образование плаценты, генерация антител и т.д.

Алексей Шацких, старший научный сотрудник лаборатории функциональной геномики Института биологии развития РАН, рассказал о том, как устроены транспозоны, могут ли эти «разрушители генов» повышать приспособленность организма, и как их можно использовать в научных исследованиях и прикладной генной инженерии.
В начале лекции докладчик напомнил основные принципы хранения и считывания наследственной информации в клетке. Гены — участки ДНК, которые кодируют белки или РНК. Эукариотические гены начинаются с промоторов и заканчиваются терминаторами, включают экзоны, входящие в зрелую мРНК, и удаляемые при ее созревании интроны. Важным этапом реализации генетической информации между стадиями транскрипции и трансляции являются различные посттранскрипционные модификации, такие как кэпирование и полиаденилирование, а также сплайсинг пре-мРНК, в ходе которого вырезаются интроны, сшиваются экзоны и образуется зрелая мРНК.
Далее лектор сделал небольшой экскурс в регуляцию транскрипции генов у эукариот. Для начала транскрипции необходимо, чтобы на промоторе образовался комплекс белков, позволяющих транскрипцию, и с промотором связалась РНК-полимераза. На инициацию транскрипции влияют энхансеры — участки ДНК, которые могут быть расположены от промотора на довольно большом расстоянии. С энхансерами связываются белки-активаторы, что приводит к образованию петли ДНК, сближающей энхансер с промотором. Этот процесс опосредует множество белков, в том числе транскрипционные факторы и белки-медиаторы. В итоге это приводит к началу синтеза РНК. И промотор, и энхансер способны регулировать уровень экспрессии генов, а также то, в каких тканях и на каких этапах они будут экспрессироваться.
Гены занимают на хромосомах фиксированное положение. Однако помимо них в геноме встречаются мобильные генетические элементы, способные к перемещению. В широком смысле к мобильным элементам относятся плазмиды, вирусы и транспозоны, но темой лекции были транспозоны, которые обычно и подразумеваются, когда речь идет о мобильных генетических элементах.
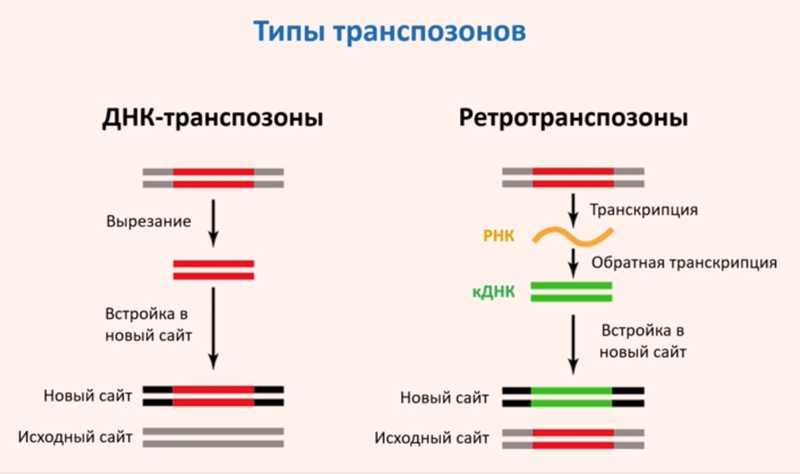
ДНК и ретротранспозоны, с LTR и без, работающие и сломанные…
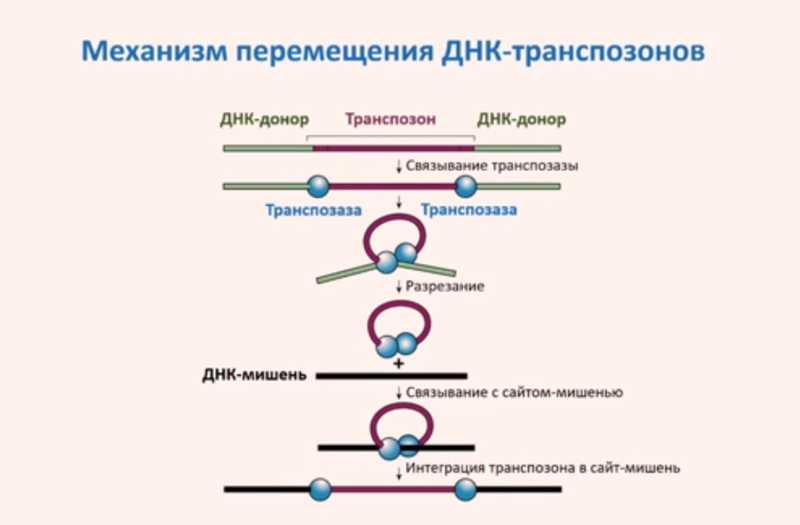
Выделяют два типа транспозонов: ДНК-транспозоны и ретротранспозоны. ДНК-транспозоны работают по принципу «вырезать-вставить», а ретротранспозоны — по принципу «копировать-вставить». ДНК-транспозоны могут перемещаться по геному за счет действия кодируемого в них белка — транспозазы, а в жизненный цикл ретротранспозонов входит стадия обратной транскрипции.
Ретротранспозоны можно разделить на те, у которых есть длинные концевые повторы (LTR) и те, у которых их нет. LTR несут в себе регуляторные элементы, обеспечивающие транскрипцию и мобилизацию транспозона. Ретротранспозоны кодируют белки с несколькими типами активности: например, с ревертазной и интегразной. Другие белки, кодируемые ретротранспозонами, могут обеспечивать формирование необходимых для их перемещения вирусоподобных частиц.
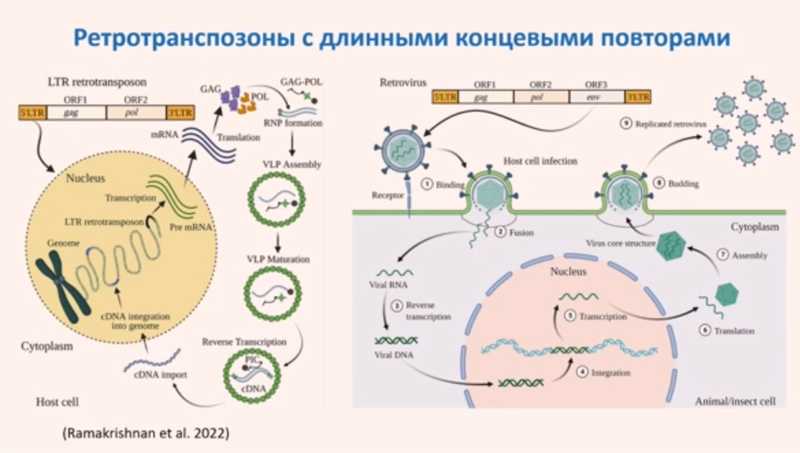
Во многом на ретротранспозоны по структуре и механизму действия похожи ретровирусы, поэтому их часто рассматривают как подтип ретротранспозонов, предполагая, что ретровирусы произошли от них в процессе эволюции. Принципиальное отличие ретровирусов от ретротранспозонов — это кодирование белка оболочки Env, с помощью которого вирусная частица может выйти за пределы клетки.
Ретротранспозоны без LTR имеют иной механизм перемещения: после транскрипции транспозона в сайт-мишень вносится одноцепочечный разрыв, к нему присоединяется РНК транспозона, при этом свободный конец ДНК используется как праймер для обратной транскрипции, а РНК — как матрица. В результате обратной транскрипции копия транспозона встраивается в новый участок ДНК.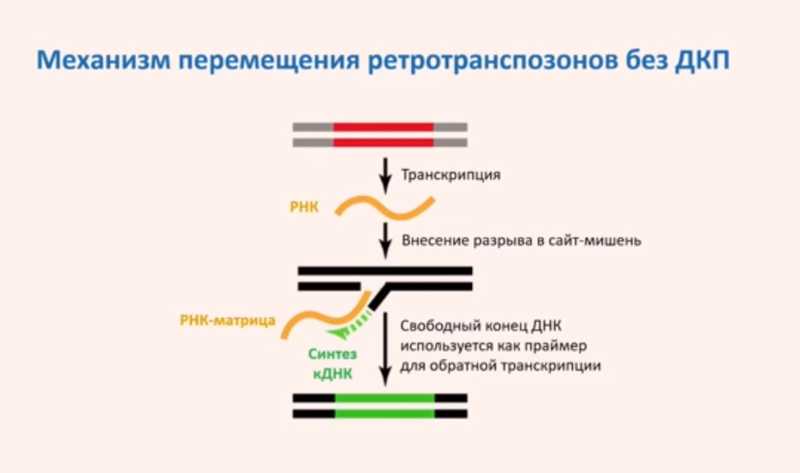
Транспозоны могут быть автономными, неавтономными и неактивными. Автономный транспозон сам кодирует все, что необходимо для его мобилизации, а неавтономный может перемещаться за счет автономных, используя их белки, обеспечивающие мобилизацию. Неактивные транспозоны появляются в ходе мутационных процессов, видоизменяющих их так сильно, что они теряют способность к мобилизации.
Вред и польза
Транспозоны составляют около 45% генома человека, хотя меньше 0,05% из них остаются активными. У растений же транспозоны могут занимать до 90% генома.
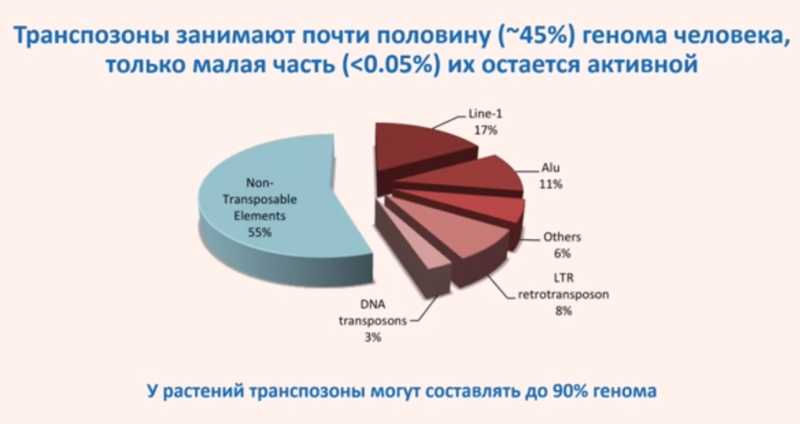
В связи с этим возникает вопрос: если транспозонов так много и они не исчезают в процессе эволюции, есть ли от них какая-то польза, приносят ли они вред или попросту бесполезны, как мусор?
Действительно, транспозоны могут быть вредны. Встраивание транспозона в ген нарушает его работу, даже если впоследствии ДНК-транспозон покинет этот ген. Также активация транспозонов может приводить к разрывам ДНК, которые могут вызывать мутации или потери участков хромосом. Даже неактивные транспозоны способны причинять вред: рекомбинация между повторяющимися копиями транспозонов может нарушить структуру хромосом, приводя к делециям или инверсиям участков.
С активностью транспозонов связаны многие патологии, такие как рак, нарушения развития, гемофилия, воспалительные и аутоиммунные заболевания, старение и ассоциированные с ним болезни, нейродегенеративные заболевания, нарушения половой системы.
В клетках существует ряд механизмов подавления активности транспозонов, благодаря которому предотвращается вред от них. Эти механизмы можно отнести к категории эпигенетических: организация хроматина и гистоновые метки, метилирование ДНК и РНК-интерференция. Так, в участках хроматина, где располагаются транспозоны, обычно присутствуют репрессивные метки на гистонах — метильные группы, из-за которых хроматин становится плотно упакованным и менее доступным для транскрипции. Метилирование ДНК тоже подавляет транскрипцию транспозонов. РНК-интерференция также может привести к подавлению транскрипции или к разрушению РНК транспозона.
В клетках зародышевого пути присутствует особый механизм РНК-интерференции — piРНК-путь. В геноме есть кластеры piРНК, в которых представлены фрагменты транспозонов. Из транскриптов этих кластеров образуются piРНК, которые распознают транспозоны и разрушают их РНК либо привлекают к ним репрессорный комплекс, способствующий эпигенетическому блокированию транскрипции транспозонов.
А есть ли от транспозонов польза? Да, и она выражается во многих аспектах. Например, транспозоны участвуют в формировании теломер и центромер хромосом. Центромеры нужны для правильного расхождения хромосом по дочерним клеткам, и в их состав входят транспозоны, обладающие определенной эпигенетической структурой. А теломераза, которая принимает участие в поддержании теломер, по всей видимости, происходит от ревертазы ретротранспозона. Кроме того, у некоторых видов сами теломеры или полностью (как у дрозофил), или частично состоят из транспозонов.
Транспозоны способствуют мутагенезу, и иногда такие мутации могут быть полезными. Например, индустриальный меланизм — изменение окраски животных из-за их обитания в зонах с высокой степенью загрязнения из-за развития промышленности — обязан своему появлению транспозонам. Березовые пяденицы Biston betularia — хрестоматийный пример индустриального меланизма: они потемнели в середине XIX века, когда стволы деревьев вокруг промышленных предприятий чернели от сажи и утрачивали лишайниковый покров, светлых особей активнее поедали птицы. Окраска пядениц меняется из-за встройки транспозона в ген cortex.

Перемещаясь по геному, транспозоны могут переносить вместе с собой регуляторные элементы и таким образом менять профиль экспрессии генов. Похожим образом транспозоны изменяют эпигенетические профили: если рядом с геном встраивается транспозон, несущий CpG-островок, который подвержен метилированию, это подавит и экспрессию соседнего гена.
В ходе ретрокопирования и неофункционализации ретротранспозоны могут создавать новые гены. Например, иногда из-за ретрокопирования в геном встраивается мРНК какого-нибудь гена, образуя псевдоген. В перспективе такой псевдоген может попасть под контроль регуляторных элементов, начать функционировать и приобрести новые свойства, став полноценным новым геном.
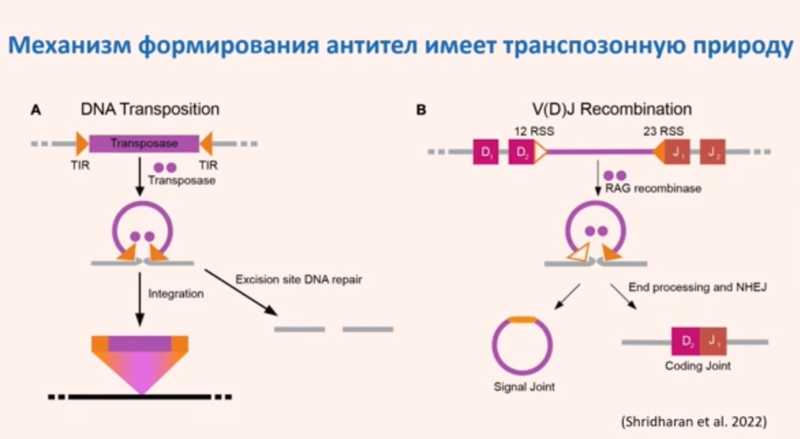
Сами по себе механизмы транспозиции, с помощью которых транспозоны перемещаются по геному, могут приобретать полезные функции. Например, механизм формирования антител — V(D)J-рекомбинация, при которой происходит рекомбинация генов иммуноглобулинов, — похож на процесс транспозиции. Транспозаза считается эволюционным предшественником RAG-рекомбиназы, которая осуществляет V(D)J-рекомбинацию.
Еще один пример «приручения» транспозона — белок Arc, который имеет ретротранспозонное происхождение и участвует в транспорте РНК между нейронами в виде вирусоподобных частиц. Arc опосредует нейропластичность и формирование долгосрочной памяти.
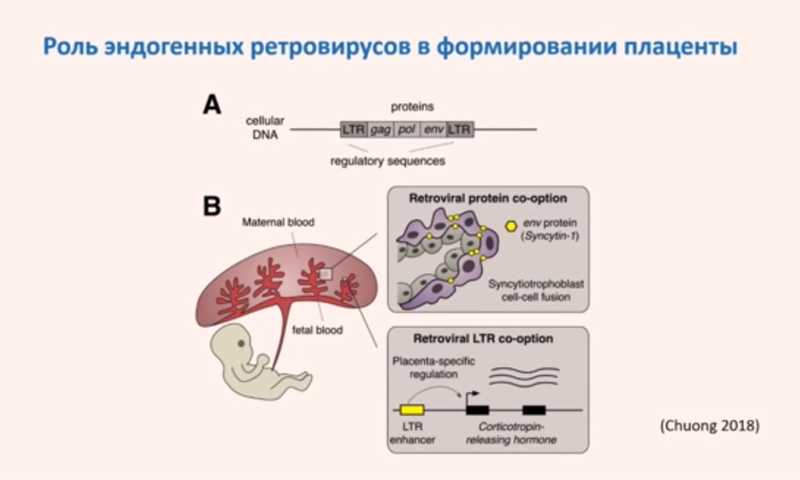
Кроме того, была показана роль эндогенных ретровирусов в формировании плаценты. Во-первых, ген env ретровируса дал начало гену Syncytin-1, который нужен для формирования синцитиотрофобласта. Во-вторых, активность гена кортиколиберина, который важен при беременности, регулируется энхансером, происходящим от LTR ретротранспозона. Благодаря такой регуляции этот ген стал экспрессироваться в плаценте, хотя ранее он экспрессировался в других тканях.
Можем ли мы использовать транспозоны?
Да — например, для изучения функций генов посредством инсерционного мутагенеза. Можно запустить в трансгенных животных или в клетках мобилизацию транспозонов, что вызовет мутации и поспособствует проявлению того или иного фенотипа. Это дает возможность отследить, какие именно мутации и гены связаны с такими фенотипами. Например, если у животного после мутагенеза появится опухоль, то можно выделить ДНК из клеток этой опухоли и выявить мутацию в гене онкосупрессора, функция которого валидируется позднее. Полученные таким образом данные помогают развивать новые методы терапии и диагностики онкологических заболеваний.
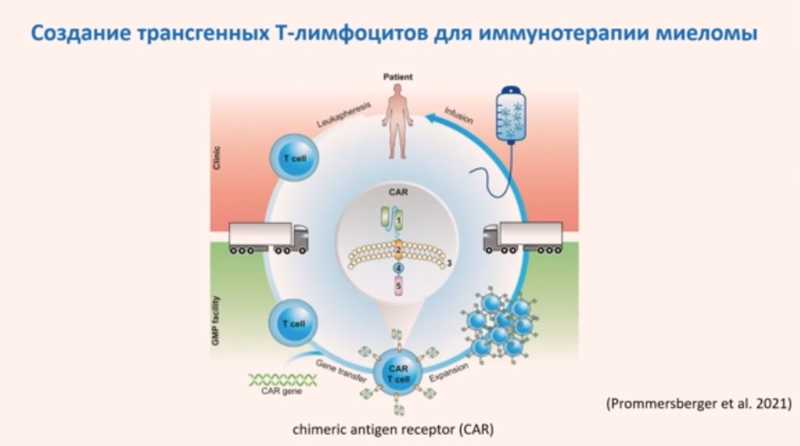
Транспозоны можно использовать как инструмент генной инженерии. Например, в клетки доставляются две плазмиды: в одной содержится интересующий ген и последовательности ДНК, которые нужны транспозону для встраивания в геном; а во второй — ген транспозазы. Когда после доставки в клетки с одной плазмиды «считается» транспозаза, она поспособствует встраиванию гена в геном клетки. Такой подход уже был использован для создания CAR Т-клеток для иммунотерапии миеломы, где транспозоны более безопасны, чем вирусные вектора.
Алексей Шацких заметил в заключение, что транспозоны оказывают огромное влияние на самые разные процессы, поэтому эта тема безгранична. По его мнению, транспозоны в каком-то смысле можно считать двигателями эволюции. Может быть, именно поэтому клетки не избавляются от них.

Вам будет интересно

 156
156
 0
0



 291
0
291
0









