
 Меню
Меню





 Все темы
Все темы
Татьяна Кочеткова: «Прокариоты — главные экстремалы планеты»
Лекция Татьяны Кочетковой, посвященная экстремофильным бактериям и археям, продолжила новый цикл научно-просветительского проекта «Метафаза» о бактериях. Она рассказала о том, как микроорганизмам удается выживать при экстремально высоких температурах, в кислоте и в соленых озерах, — и, конечно, о том, как такие микроорганизмы помогли развитию молекулярной биологии и биотехнологии.

В продолжение цикла научно-просветительского проекта «Метафаза», посвященного бактериям, Татьяна Кочеткова (к.б.н., старший научный сотрудник отдела биологии экстремофильных микроорганизмов ФИЦ «Фундаментальные основы биотехнологии» РАН) выступила с лекцией «Жизнь на грани фантастики: почему микробы — главные экстремалы планеты».
«Нормальные условия» для различных организмов — оптимальные для их жизнедеятельности температура, pH, соленость, наличие кислорода и питательных веществ — варьируют очень сильно. Для мезофилов, в число которых можно было бы включить и людей (хотя этот термин применим все же к микроорганизмам), нормальные условия — температурный диапазон в 20–45 °С и нейтральный pH. Однако в микромире есть и экстремофильные организмы, которые с нашей точки зрения живут в необычных условиях, в «крайностях», которые для них нормальны.
Перед тем как перейти к разговору об экстремофилах, Татьяна Кочеткова рассказала об археях. В 1990 году Карл Вёзе выделил их в третий домен живых организмов наряду с бактериями и эукариотами, построив филогенетическое дерево организмов по последовательности генов их 16S рРНК (до Вёзе они считались подгруппой бактерий). Более новые исследования, однако, склоняются к двухдоменной системе: изначально существовали бактерии и археи, а плод их «сотрудничества» дал начало эукариотам.
Археи, как и бактерии, являются одноклеточными прокариотами. Они похожи на них внешне и имеют метаболические сходства. И все же археям свойственны особые молекулярные механизмы, связанные с сохранением и передачей информации, делением и цитоскелетом, более близкие к аналогичным процессам в эукариотических клетках, чем в бактериальных. Также у архей есть уникальные типы метаболизма (например, метаногенез), среди них не выявлено патогенов и на них не действуют антибиотики.
Жители горячих источников
Среди прокариот встречаются термофилы — для таких микроорганизмов комфортными являются температуры выше 45 °С вплоть до 122 °С. Обитают термофилы в горячих источниках, как наземных, так и глубоководных («черные» и «белые курильщики»). Они могут найти себе жилье в горячих компостных кучах и геоэлектростанциях.
Как правило, термофилы живут в анаэробных условиях. В их местах обитания присутствуют вулканические газы — водород, сероводород, угарный газ, метан. Эти восстановленные неорганические соединения обладают большим количеством потенциальной энергии и способны быть донорами электронов, что термофилы используют в своем метаболизме. Также в среде обитания термофилов могут в больших количествах содержаться тяжелые металлы и токсичные вещества (мышьяк, ртуть).
Так как вулканические газы разнообразны по своему составу, термофилы характеризуются огромным спектром метаболических возможностей. Чаще всего в качестве донора электронов они используют водород и часто являются автотрофами, т.е. организмами, которые образуют органические вещества из неорганических самостоятельно.
На данный момент организмом, который живет при наиболее высоких температурах, является Methanopyrus kandleri, он выдерживает температуры до 122 °С. Для получения энергии эта архея проводит метаногенез. А наименее энергоемкую реакцию «открыл» Thermococcus onnurineus, который превращает формиат в водород и углекислый газ. Особенности его метаболизма были описаны в журнале Nature совместно корейскими и российскими учеными в 2010 году.

В начале 2000-х гг. ученые обнаружили в глубоководных «курильщиках» необычного симбионта гипертермофильной археи Ignicoccus hospitalis — «наноархею» Nanoarchaeum equitans. Размер их клеток — 400 нм, а размер генома — всего лишь половина мегабазы, этом самый маленький известный геном на данный момент.
Позднее биоинформатический анализ показал, что термофильные и гипертермофильные бактерии и археи находятся у самого корня филогенетического дерева всех организмов. Именно эти микроорганизмы, возможно, были первыми обитателями Земли.
Хотя большая часть гипертермофилов представлена археями, известны и бактерии, живущие при высоких температурах. Две самые высокотемпературные бактерии — Thermotoga maritima (оптимум 80 °С) и Aquifex pyrophilus (опт. 85 °С).
Что термофилы дали молекулярным биологам
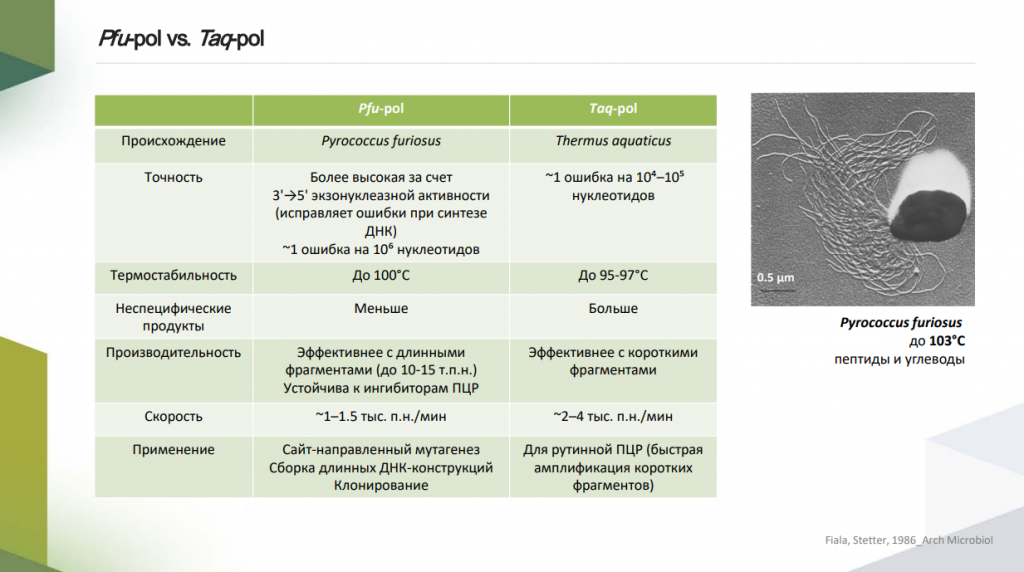
Еще одна термофильная бактерия, которая может жить в диапазоне температур 40–79 °С, — Thermus aquaticus — особенно значима с точки зрения биотехнологии. Она была выделена из Йеллоустонского горячего источника. Что интересно, она стала первым культивируемым термофильным микроорганизмом. Эта бактерия помогла развитию технологии полимеразной цепной реакции (ПЦР), ведь из Thermus aquaticus была выделена термостабильная Taq-полимераза.
До того, как в обиход вошла Taq-полимераза, для синтеза цепей ДНК использовалась обычная полимераза из E. coli. Так как для денатурации ДНК, отжига праймеров и элонгации требуются различные температуры, реакционную смесь нужно было нагревать, при этом полимераза E. coli денатурировала, и ее приходилось добавлять снова и снова в каждом цикле. Чтобы обойти эту трудность, американский биохимик Кэрри Муллис — создатель ПЦР — предложил использовать Taq-полимеразу, способную работать при высоких температурах без снижения активности.
В молекулярной биологии популярна и другая термостабильная полимераза — Pfu-полимераза, выделенная из Pyrococcus furiosus. Эта архея может жить при температурах до 103 °С. По сравнению с Taq-полимеразой Pfu более термостабильна и точна, однако отличается меньшей процессивностью. Если Taq применяют в рутинной ПЦР, то Pfu используется для более сложных процедур: клонирования, сборки длинных ДНК-конструкций, сайт-направленного мутагенеза.
Секреты термоустойчивости
Как же термофилам удается выживать при высоких температурах? Для белков термофилов характерна особая вторичная и третичная структура: гидрофобные аминокислотные остатки они «прячут» внутрь молекулы, а гидрофильные, наоборот, остаются снаружи и притягивают воду. В этих белках также повышено число ионных и водородных связей, дисульфидных мостиков. Все это обеспечивает плотную укладку белковых молекул, которая делает их устойчивыми ко многим воздействиям.
Для термофилов и гипертермофилов также характерен маленький размер генома. Благодаря этому они тратят меньше ресурсов на репарацию своей ДНК.
Кроме того, в клетках термофилов содержатся уникальные полиамины, имеющие длинные либо разветвленные цепочки. Их роль состоит в стабилизации ДНК и РНК при повышенных температурах. Гипертермофилы, у которых отсутствует фермент для синтеза таких полиаминов, не способны жить при температурах выше 90 °С.
Многие термофилы имеют особый белок — обратную гиразу. При высоких температурах двойная цепочка ДНК расплетается, что препятствует работе ферментов, взаимодействующих с ней. Обратная гираза обращает денатурацию ДНК вспять, восстанавливая ее двухцепочечную структуру. Считается, что впервые обратная гираза появилась у архей, а потом ее «позаимствовали» вместе с другими генами термофильности бактерии — уже упоминавшиеся Thermotoga maritima и Aquifex pyrophilus. А позднее они «раздали» способность жить при высоких температурах другим видам бактерий с помощью горизонтального переноса генов.
Наконец, термофильные археи имеют особые мембраны. В их состав входят фосфолипиды, содержащие вместо жирных кислот спирты, простые эфирные связи вместо сложноэфирных и имеющие особую пространственную конфигурацию.
Прокариоты, которым не страшны кислоты
Прокариоты могут жить в кислотах — при pH < 5. Наибольшее разнообразие ацидофилов также характерно для архей. Среди ацидофилов можно выделить термоацидофилов, которые живут в кислых горячих источниках. Считается, что эти источники сформировались под влиянием термофилов после «кислородного взрыва», который случился около 2,4 млрд лет назад. После того как цианобактерии научились вырабатывать кислород, другие микроорганизмы начали использовать кислород в своем метаболизме. Он оказался удобным, наиболее энергетически выгодным акцептором электронов. Термофилы тоже стали использовать кислород, при этом окисляя различные соединения серы из горных пород и вулканических газов. Таким образом в горячих источниках начала накапливаться серная кислота.
Термоацидофилы, в основном представленные археями, — самая многочисленная группа ацидофилов. К ним относятся Sulfolobus sp. (pH 2,0–3,0, 75–80 °С), ‘Tardisphaera miroshnichenkoae’ (pH 4,0, 55 °С), Acidilobus aceticus (pH 3,5–4,0, 85 °С), Picrophilus oshimae (pH 0–3,5, 60 °С). Последняя архея — рекордсменка по жизни в самых кислых условиях.

Для выживания в кислоте ацидофилы «научились» создавать обратный мембранный потенциал. У других клеток отрицательный заряд сосредоточен с внутренней стороны мембраны, а положительный — с внешней. Это нужно для работы АТФаз, использующих протоны и вырабатывающих энергию. Чтобы обезопасить себя от проникновения в клетку протонов, ацидофилы активно закачивают внутрь клетки ионы калия.
Мембраны ацидофильных архей малопроницаемы для протонов и сами по себе. В их состав входят тетраэфирные липиды из изопреноидных остатков. Некоторые бактерии позаимствовали эту стратегию и тоже делают тетраэфирные липиды из остатков жирных кислот.
Если же протоны кислот все-таки проникли в клетку, ацидофилы их выкачивают с помощью разных транспортеров. Обычно для работы таких транспортеров требуется тратить энергию в виде АТФ, но ацидофилы преимущественно применяют вторичные транспортеры, которые работают за счет разности потенциалов. Внутри клетки протоны нейтрализуются также с помощью «буферных» молекул: полифосфатов, большого количества остатков гистидина. Денатурации белков препятствуют шапероны и термосомы (шапероны архей), а протеасомы обеспечивают утилизацию необратимо поврежденных белковых молекул.
Ацидофилы в техногенных нишах
Ацидофилы живут и в относительно холодных условиях, причем часто — в техногенных местах обитания: шахтных дренажах, затопленных карьерах горнодобывающей промышленности. В такой среде уровень pH может опускаться до отрицательных значений, в ней также содержатся большие количества сульфатов и металлов. К ацидофилам, живущих в техногенных локациях, относят рода Acidithiobacillus (pH 1,5–2,5), Leptospirillum (pH 1,0–2,0), Ferroplasma (pH 1,7).
Ацидофилы — обитатели техногенных ниш в основном окисляют железо и серу. Этот естественный процесс приводит к экологическим проблемам: из-за неосторожного использования горных ресурсов кислая вода попадает в обычные водоемы, приводя к гибели живущих в них организмов.
Те же микроорганизмы используются человеком в биогидрометаллургии: процесс биовыщелачивания помогает более эффективно и экологично разрабатывать железо-серные руды, а также очищать стоковые воды от ядовитых соединений и сульфатов.
Жизнь в Мертвом море
Еще одна группа экстремофильных прокариот — галофилы — предпочитают среды с концентрацией соли 30–300 г/л. Основные места обитания этих экстремофилов — гиперсоленые озера, такие как Мертвое море, и даже подводные соленые брины. В зависимости от происхождения, в таких водоемах наблюдают высокие уровни ионов кальция, магния, хлоридов. В местах обитания с высоким содержанием соды (2–5 М) обитают галонатронофилы. Помимо вышеперечисленного, в таких водоемах низкаяактивность воды, то есть в них содержится мало молекул воды, которые могут быть использованы для метаболизма. Примеры галофилов — Halobacterium salinarum (это первый выделенный галофил), Haloferax volcanii (часто используется в молекулярной биологии для трансформации), Haloarcula japonica (имеет треугольные клетки), Salinibacter ruber (одна из немногих экстремально галофильных бактерий).
Еще один интересный представитель галофилов — архея Haloquadratum walsbyi. Она живет в солнечных соляных прудах, а клетки этой археи имеют форму квадрата и достигают 40 мкм. Эта архея умеет создавать вакуоли, чтобы подниматься к поверхности водоема для контакта с кислородом. Она также считается рекордсменом по жизни в очень сухих условиях, где активность воды составляет всего лишь 0,63. Еще эта архея выделяет белок галомуцин, похожий на слизь: благодаря ему она сохраняет вокруг себя оболочку из молекул воды. На рисунке ниже (в левом нижнем углу) галомуцин окрашен зеленым, а сама архея — красным. Что интересно, галомуцин — очень крупный белок, для синтеза и выкачивания его наружу из клетки теоретически требуется около 180 молекул АТФ.

У галофилов есть две стратегии выживания — salt-in и salt-out. Первую, менее энергозатратную, используют экстремальные галофилы (археи и некоторые бактерии): они создают внутри клетки высокие концентрации ионов калия, взаимодействующих со всеми внутриклеточными белками, которые в других организмах используют ионы натрия.
Умеренные галофилы (бактерии, водоросли, грибы и некоторые археи) используют вторую, более энергозатратную стратегию: накапливают защитные вещества, такие как эктоин, бетаин, трегалоза и т.д. В этом случае микроорганизмы расходуют энергию, чтобы «затащить» эти крупные молекулы внутрь клетки или синтезировать их самостоятельно.
Археи, которые используют стратегию salt-in, поддерживают мембранный потенциал также за счет выкачивания протонов из клетки и закачивания хлорид-анионов с помощью бактериородопсина и галородопсина соответственно. Для работы этим белкам требуется солнечный свет.
Пурпурные мембраны
Бактериородопсин был впервые выделен из Halobacterium salinarum в 1970 году. Свое название он получил из-за сходства с родопсином, который присутствует в сетчатке глаза млекопитающих. Бактериородопсин способен переносить протоны через мембрану против электрохимического потенциала и преобразует энергию света (с длиной волны 500–650 нм) в химическую за счет сопряжения с АТФазами. При этом бактериородопсин характеризуется высокой стабильностью при высокой солености (до 5 М), температуре (до 140 °С) и различных pH (5,0–11,0).
Прокариоты, несущие в мембране бактериородопсины, имеют пурпурные мембраны. Именно это, в добавлении к каротиноидам галофилов, и обуславливает характерный розовый цвет соленых озер.
Бактериородопсин имеет большое значение для фундаментальной и прикладной науки. Его можно использовать в качестве модели GPCR (рецепторов, сопряженных с G-белком — семиспиральным рецепторам, к которым относится большая часть известных лекарственных мишеней у человека) и мембранных транспортеров. Бактериородопсин также применяется для разработки новых носителей информации, для создания искусственной сетчатки и в составе оптических сенсоров.
Таким образом, изучение биоразнообразия экстремофильных прокариот помогает лучше понять эволюцию организмов, исследовать грани метаболических возможностей и даже выстраивать гипотезы о жизни на других планетах, где условия отличаются от земных. И, конечно, благодаря экстремофилам биотехнологи приобретают все новые удобные инструменты.
Другие лекции блока «Бактерии и археи»
Матвей Колесник о гонке вооружений между бактериями и фагами
Евгений Фролов: «Метаболизм ацетогенов — это конструктор»
Павел Сольев: «Подавлять ферменты бактерий нужно с умом»

Вам будет интересно

 156
156
 0
0



 291
0
291
0









