Меню
Меню





 Все темы
Все темы
Длина тонкого кишечника и его ворсинок увеличивается при беременности у мышей
Международная группа исследователей продемонстрировала, что длина и масса тонкого кишечника повышаются уже на седьмой день беременности у мышей, а к концу беременности прирост длины достигает 18%. После окончания вскармливания кишечник не возвращается к изначальному размеру, в отличие от ворсинок, которые сначала удлиняются, а потом укорачиваются. Эффект связан с активностью транспортера глюкозы SGLT3a, чувствительного к изменению pH и уровня натрия. Удлинения ворсинок тонкого кишечника можно добиться повышением содержания натрия в диете мыши.
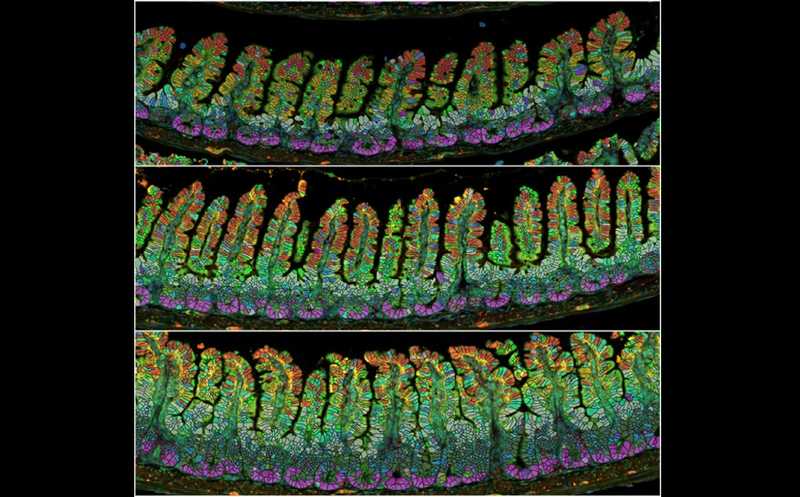
Эпителий кишечника девственных (вверху), беременных (в центре) и кормящих (внизу) мышей.
Credit:
Tomotsune Ameku | Пресс-релиз
Кишечник отличается повышенной пластичностью: его эпителий постоянно самообновляется. В тонком кишечнике пролиферирующие клетки крипт дают начало клеткам ворсинок, которые мигрируют и дифференцируются в энтероциты и секреторные клетки, прежде чем пройти через апоптоз на кончике ворсинки. Однако пластичность кишечника в основном изучается при повреждении или недостатке нутриентов и микроорганизмов, а ведь на органы влияет еще и репродукция. Ранее международная группа ученых показала, что кишечник самки дрозофилы увеличивается при появлении потомства. Однако механизм этого процесса плохо изучен. В новой работе та же группа исследователей изучила механизмы увеличения размера кишечника у мышей при беременности и лактации.
У самцов мышей и девственных самок размеры тонкого кишечника сопоставимы. Но в период беременности и лактации длина и масса кишечника у самок значительно повышаются. Это становится заметным уже на седьмой день беременности, а к 18-му дню тонкий кишечник уже на 18% длиннее, чем у девственных самок. Этот процесс обратим, но не до конца: даже спустя 35 дней после окончания лактации у рожавших самок тонкий кишечник был длиннее. А в ходе второй беременности он удлиняется еще сильнее.
Гистология показала, что при беременности крипты кишечника становятся глубже, а ворсинки — шире и на 20–30% длиннее. Эти изменения тоже заметны на седьмой день беременности, однако уже через семь дней после окончания лактации они возвращаются в изначальное состояние. И с последующими беременностями изменения не накапливаются. В дальнейших экспериментах изучался период временем между седьмым днем беременности и седьмым днем лактации.
В рассматриваемый период повышалось количество пролиферирующих клеток в тонком кишечнике, особенно в криптах. Также активировалась миграция клеток из крипт в ворсинки. Однако количество клеток, проходящих через апоптоз, не снижалось. В то же время размеры дифференцирующихся эпителиальных клеток в ворсинках увеличивались. Этот эффект был также обратимым. Таким образом, рост ворсинок связан с повышением темпов пролиферации клеток-предшественников, скорости миграции и размеров эпителиальных клеток.
Авторы ответили на вопрос, связан ли рост кишечника с повышением объемов потребления пищи в период беременности и лактации. Увеличение размера кишечника заметно уже на седьмой день беременности, когда мышь еще не начинает усиленно питаться (это происходит на 12–13 день).
Беременность влияет на микробиом кишечника. Также известно, что у мышей без микробиома кишечник длиннее. Авторы предположили, что изменения, наблюдаемые при беременности, ассоциированы в том числе с микробиомом. Однако у мышей, выращенных в стерильных условиях, тонкий кишечник все равно удлиняется при беременности и лактации, а крипты и ворсинки претерпевают те же изменения, что и у мышей с нормальным микробиомом. Те же адаптации были показаны у мышей, получавшие пробиотик (Lactiplantibacillus plantarum) в ходе беременности.
РНК-секвенирование продемонстрировало значительные изменения транскрипции в кишечнике у беременных мышей. Изменения были очевидны в двенадцатиперстной, тощей и подвздошной кишке и включали гены, участвующие в метаболизме, сигналинге и иммунитете.
Авторы провели РНК-секвенирование единичных клеток в подвздошной кишке, так как там изменения транскриптома были наиболее выраженными. Ремоделирование, связанное с беременностью и лактацией, было сильнее всего заметно в энтероцитах. У кормящих самок на более высоком уровне экспрессировались гены, связанные с метаболизмом жирных кислот, ацил-кофермента А и пирувата, транспортом липидов, желчных кислот, сахара и микронутриентов. Многие наблюдения подтвердили методом smFISH (single-molecule fluorescence in situ hybridization).
Беременность и лактация повышали уровень экспрессии Slc5a4a в эпителии кишечника. Slc5a4a кодирует белок SGLT3a, член семейства натрий-глюкозных котранспортеров и один из двух паралогов у мышей (SGLT3a и SGLT3b). Slc5a4a экспрессируется в энтероцитах у основания и в середине ворсинки.
SGLT3a отвечает за активный транспорт глюкозы через мембрану. Авторы провели нокаут белков SGLT3 или SGLT3a. Фенотип мышей с нокаутом внешне не изменился, однако мыши с нокаутом SGLT3 хуже набирали вес при вскармливании. Также нокаут сильно повлиял на многие процессы, связанные с метаболизмом и транспортом различных веществ. Авторы показали, что SGLT3a чувствителен к pH и натрию. Повышение уровня натрия в диете мышей (но не кислоты) способствовало пролиферации эпителия и росту ворсинок. У самок с нокаутом SGLT3 длина ворсинок при беременности увеличивалась не так значительно, как у мышей дикого типа, но на длину тонкого кишечника нокаут не влиял. Дополнительные эксперименты подтвердили, что SGLT3a необходим для более интенсивной пролиферации эпителия тонкого кишечника, связанной с беременностью и лактацией.
В настоящее время авторы изучают, подвергаются ли другие типы клеток в кишечнике мышей ремоделированию во время беременности, и происходит ли это у людей.
Источник:
Tomotsune Ameku, et al. Growth of the maternal intestine during reproduction // Cell (2025), published 19 March 2025, DOI: 10.1016/j.cell.2025.02.015

Вам будет интересно

 192
192
 0
0