Меню
Меню





 Все темы
Все темы
Для образования внеклеточных везикул архее необходима малая ГТФаза
Археи, подобно бактериям и эукариотам, могут секретировать внеклеточные везикулы. Авторы статьи, опубликованной в PNAS, показали, что архее Haloferax volcanii для этого необходима малая ГТФаза, которую они назвали ArvA. Внеклеточные везикулы H. volcanii содержат разнообразные РНК и могут поглощаться другими клетками. Авторы выдвинули предположение об эволюционных связях между продукцией везикул у архей и внутриклеточным везикулярным транспортом у эукариот, в котором также участвуют ГТФазы.
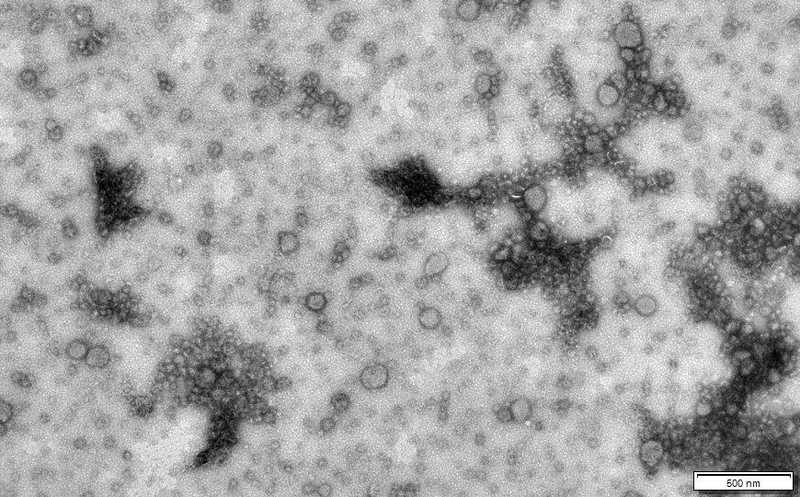
Везикулы Haloferax volcanii
Credit:
S. Erdmann/Max Planck Institute for Marine Microbiology | Пресс-релиз
Внеклеточные везикулы — небольшие мембранные пузырьки, которые отпочковываются от клеточной мембраны. Их способны секретировать клетки всех трех доменов живых организмов. Везикулы могут содержать белки, нуклеиновые кислоты, сигнальные молекулы и участвовать в межклеточной коммуникации. Достаточно много известно про везикулы, секретируемые бактериями, а про везикулы архей — гораздо меньше. Однако установлено, что некоторые представители типа Euryarchaeota продуцируют везикулы, содержащие плазмидную ДНК и, возможно, таким путем у архей происходит горизонтальный перенос генов. Авторы статьи, опубликованной в PNAS, на примере археи Haloferax volcanii установили, что эвриархеотам для образования внеклеточных везикул необходима малая ГТФаза.
Ранее было показано, что у галоархеи H. volcanii — экстремофила, живущего в соленой воде, — внеклеточные везикулы образуются при УФ-облучении. Авторы обсуждаемой работы собрали везикулы из супернатанта культуры H. volcanii. Они представляли собой сферические тельца размером 50–150 нм в диаметре. Когда H. volcanii культивировали при оптимальной для нее температуре 45 °C, везикул в супернатанте было мало, однако при понижении температуры до 28 °C количество везикул увеличивалось в 2,7 раза. Повышенное образование внеклеточных везикул наблюдалось и при других видах стресса, таких как облучение ультрафиолетом и вирусная инфекция.
Исследователи попытались выделить из внеклеточных везикул H. volcanii нуклеиновые кислоты, и оказалось, что они заполнены РНК. Содержание рРНК в них было примерно таким же, как в клетке, содержание коротких транскриптов было повышено. Дальнейшие исследования показали, что внеклеточные везикулы H. volcanii содержат также тРНК и другие некодирующие РНК.
РНК в составе везикул представляет транскрипты примерно 4400 генов H. volcanii — это более трех четвертей всего генома. Впрочем, большую часть этих РНК следует рассматривать как транскрипционный шум в силу их очень малого количества. Больше всего в везикулах тРНК, потом следуют некодирующие РНК (все РНК, кроме мРНК, тРНК и рРНК), на третьем месте рРНК. Матричные РНК поступают во внеклеточные везикулы преимущественно в виде фрагментов транскриптов.
Самые многочисленными среди некодирующих РНК оказались межгенные смысловые и антисмысловые РНК. Функции их неизвестны, кроме того, везикулярные некодирующие РНК не содержат каких-либо консенсусных последовательностей или вторичных структур, которые можно было бы рассматривать как сигналы для упаковки в везикулы. Интересно, что внеклеточные везикулы, отпочковавшиеся от клеток, зараженных вирусом, никак не отличаются от везикул, произведенных неинфицированными клетками. Образование внеклеточных везикул, наполненных РНК, было показано и для других галоархей.
В состав внеклеточных везикул H. volcanii входят и белки. Особенно многочисленны белки группы CetZ, которые контролируют форму и подвижность клеток, а также FtsZ2, контролирующий клеточное деление, АТФ-связывающий белок ABC-транспортера, пептидаза сигнального пептида и несколько гипотетических белков. По составу мембранных белков внеклеточные везикулы отличаются от клеточной мембраны. Например, в мембранах везикул, отпочковавшихся после обработки ультрафиолетом, отсутствует малая ГТФаза HVO_3014. Внеклеточные везикулы также отличаются повышенным содержанием диаденилатциклазы, которая образует циклический диАМФ — распространенный вторичный посредник у бактерий и архей.
Ключевую роль в формировании заполненных РНК внеклеточных везикул у H. volcanii играет малая ГТФаза OapA. Ее нокаут делает невозможным образование таких везикул, хотя клетки с нокаутом по OapA росли чуть быстрее, чем клетки дикого типа. Авторы исследования предложили для OapA название ArvA — Archaeal vesiculating GTPase, ГТФаза архей, необходимая для образования везикул.
У других групп архей присутствуют гомологи ArvA, обозначенные ArvB и ArvC. В то же время белок ArvA консервативен среди многих клад архей. Авторы высказали предположение, что ГТФазы, влияющие на везикулярный транспорт, возникли в ходе эволюции достаточно рано. Некоторые ГТФазы бактерий и архей кластеризуются с эукариотическими ГТФазами, и можно предположить, что механизмы внутриклеточного везикулярного транспорта эукариот (в котором ГТФазы также играют важную роль) унаследованы не от бактериальных эндосимбионтов, а от архей.
Дальнейшие исследования показали, что клетки H. volcanii захватывают РНК из везикул, причем обмен РНК между клетками с помощью везикул происходит довольно быстро. Это подтверждает, что везикулы могут служить для межклеточной передачи сигналов.
Источник
Joshua Mills, et al. Extracellular vesicle formation in Euryarchaeota is driven by a small GTPase // PNAS, published February 26, 2024, 121 (10), e2311321121, DOI: 10.1073/pnas.2311321121




 0
0