Меню
Меню





 Все темы
Все темы
Гематоэнцефалический барьер формируется при участии «временно состаренных» клеток
Гематоэнцефалический барьер (ГЭБ) — избирательный физиологический «фильтр» между центральной нервной системой и кровотоком. Несмотря на важную роль этой структуры, механизм ее эмбрионального развития не был ясен до конца. Авторы статьи в Cell показали на эмбрионах мышей, что в нем участвуют сенесцентные клетки, причем такой фенотип они приобретают временно. Маркер сенесценции p21 экспрессировался в резидентных макрофагах, клетках эндотелия сосудов, а также в сосудистом сплетении, которое участвует в выработке спинномозговой жидкости (СМЖ). Сенесценция в них ограничивала ангиогенез и поддерживала эпителиальную функцию. Удаление этих клеток приводило к избыточному росту сосудов и снижало выработку СМЖ, что в итоге нарушало нормальную функцию ГЭБ.
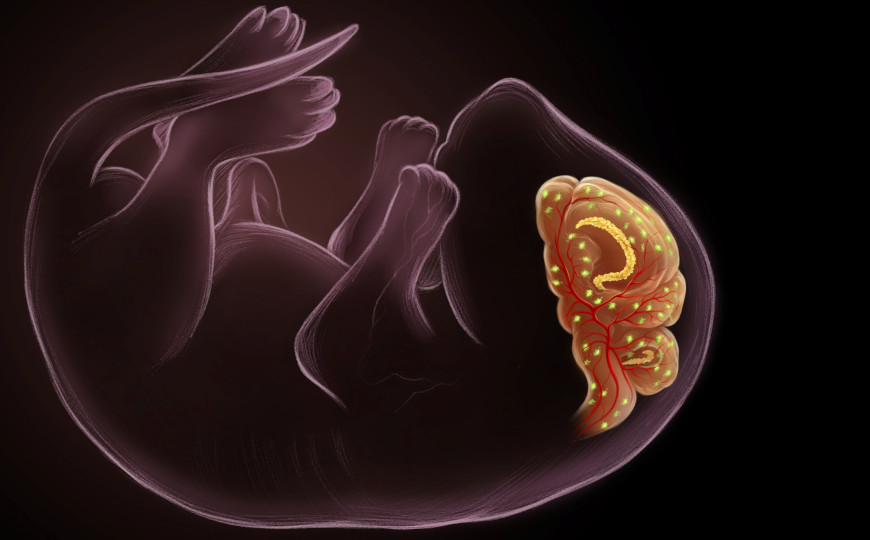
Ella Maru Studio, conceptualized by Ashley Watson and Hiruy Meharena | пресс-релиз
Гематоэнцефалический барьер (ГЭБ) — одна из важнейших защитных структур организма. Его специализированные клетки пропускают в мозг питательные вещества, но препятствуют попаданию токсинов и патогенов из кровотока. Формирование ГЭБ в ходе развития изучено недостаточно. Его проанализировали авторы статьи в Cell, которые показали на мышах, что важную роль в этом процессе играют сенесцентные клетки.
Уже известно, что повышенная экспрессия p21 — это характерный признак сенесценции, в том числе запрограммированной сенесценции при эмбриогенезе. Ученые отследили экспрессию p21 в тканях мышиных эмбрионов. На 13,5 сутки эмбрионального развития (E13.5) p21+ клетки в большом количестве обнаружились в сосудистом сплетении (ChP) боковых и четвертого желудочков мозга, а также в кортикальном крае — временной структуре на границе между гиппокампом и передним мозгом.
Паттерны экспрессии во времени отличались между этими областями. В кортикальном крае количество p21+ клеток достигало пика на E12.5–E13.5 и исчезало к E18.5. Напротив, в желудочках мозга оно достигало пика на E13.5–E14.5 и оставалось повышенным.
Затем ученые проанализировали опубликованные данные секвенирования РНК единичных клеток, чтобы выявить конкретные типы p21+ клеток на этом этапе эмбриогенеза. Ими оказались ранние нейроны (клетки Кахаля — Ретциуса), клетки ChP, эндотелиоциты сосудов (VEC) и резидентные макрофаги. Авторы экспериментально подтвердили экспрессию ключевых маркеров этих линий в p21+ клетках.
Анализ транскриптома показал, что все p21+ клетки эмбрионального мозга, кроме клеток Кахаля — Ретциуса (CR), обладают сенесцентным фенотипом. В них была снижена экспрессия генов, связанных с клеточным циклом, репликацией ДНК и трансляцией белков, и повышена — генов генов воспалительной реакции, функции лизосом, продукции цитокинов, катаболизма липидов и везикулярного транспорта. Повреждения ДНК или апоптоза в них не наблюдалось. В CR ученые не нашли выраженных признаков сенесценции, поэтому предположили, что в этих клетках транзиторная экспрессия p21 играет роль скорее в дифференцировке.
В ходе эмбриогенеза p21+ клетки ChP, VEC и резидентные макрофаги не входили в клеточный цикл, сохранялись по крайней мере до трех недель после рождения. Однако в постнатальном периоде они переставали экспрессировать маркеры сенесценции.
Более подробный анализ транскриптома каждой из p21+ субпопуляций указал на их роль в регуляции ангиогенеза. Авторы подтвердили это, удалив p21+ клетки из эмбрионального мозга мыши, что привело к увеличению площади и ветвления сосудов. Селективное истощение p21+ макрофагов имело обратный эффект, а удаление p21+ клеток ChP нарушало функцию эпителиальной ткани.
Таким образом, взаимодействие нескольких популяций сенесцентных клеток в головном мозге необходимо для нормального развития ГЭБ. Совместно эти клетки направляют ангиогенез и одновременно обеспечивают барьерную функцию, чтобы сформировать этот сложный «фильтр» между кровотоком и центральной нервной системой.
Метилирование мРНК регулирует формирование переднего мозга у мышей
Источник
Watson L, et al. Persistent and transient senescent cells contribute to brain-barrier development. // Cell, 2026. DOI:
10.1016/j.cell.2026.05.022

Вам будет интересно



 144
144
 0
0