Меню
Меню





 Все темы
Все темы
Пролактин и окситоцин активируют «контур агрессии» в мозге кормящей мыши
Авторы статьи в Nature Communications с помощью опытов на мышах описали механизм, который позволяет обычно спокойным самкам млекопитающих внезапно переходить к яростной защите своих детенышей. Ученые обнаружили нейронный контур, который отвечает за материнскую агрессию (и за агрессию в целом), однако в норме активен только у самцов, а в мозге нерожавших самок пребывает в «спящем состоянии». Этот контур временно активируется материнскими гормонами (пролактином и окситоцином), давая доступ к универсальной поведенческой программе агрессии.
Агрессивное поведение, как правило, зависит от пола — у многих видов, включая лабораторных мышей, она ярко выражена у самцов, тогда как самки вне послеродового состояния не агрессивны. Однако после родов и во время лактации самки проявляют материнскую агрессию — резкое, но обратимое изменение социального поведения. Авторы статьи в Nature Communications проанализировали это явление, чтобы понять, зависят ли поведенческие программы, связанные с полом, от принципиально разных (диморфных) или общих (мономорфных) нейронных цепей.
Несмотря на то, что роль материнских гормонов в заботе о потомстве хорошо известна, их влияние на материнскую агрессию, особенно на клеточном уровне, оставалось неясным. Более того, было неизвестно, являются ли нейронные модули, вызывающие агрессию, общими для обоих полов.
В поисках ответа ученые сосредоточились на вентральном премаммиллярном ядре (PMv) гипоталамуса. Это ядро известно как часть «области атаки» в гипоталамусе, уже показано, что оно задействовано в агрессии между самцами. В частности, внимание авторов привлекли глутаматергические нейроны, экспрессирующие ген дофаминового транспортера (PMvDAT).
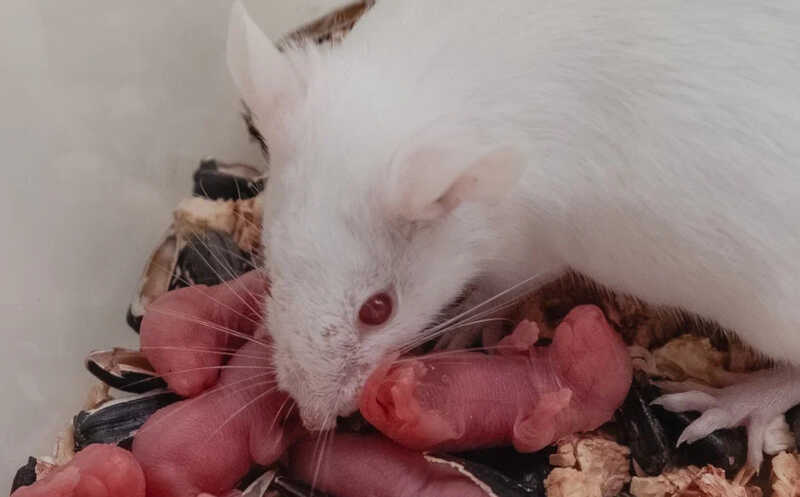
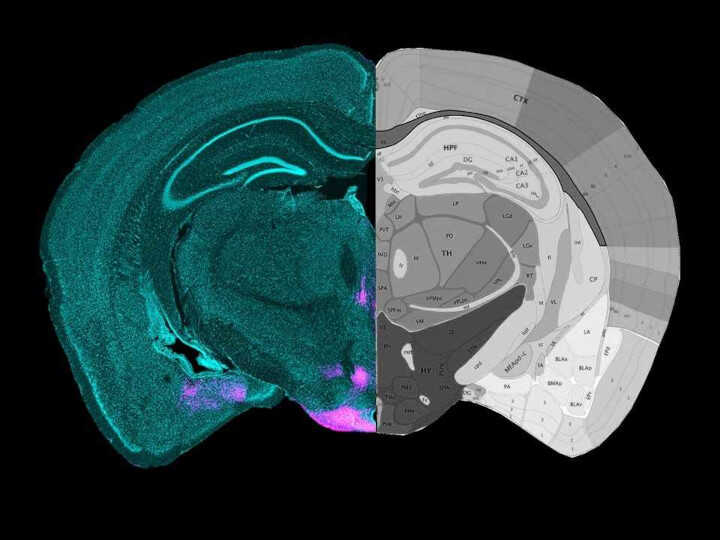 Розовым окрашены нейроны вентрального премаммиллярного ядра гипоталамуса мыши.
Розовым окрашены нейроны вентрального премаммиллярного ядра гипоталамуса мыши.Credit:
Broberger Lab | пресс-релиз
Чтобы проверить, участвуют ли эти нейроны в материнской агрессии, исследователи проанализировали агрессивных кормящих самок, используя парадигму «резидент-нарушитель» (Resident-intruder, RI). Около 42% самок демонстрировали выраженную агрессию к нерожавшей самке на своей территории, схожую по своим паттернам с агрессией самцов.
Регистрируя электрическую активность живых срезов мозга методом пэтч-кламп, авторы установили ключевое различие. PMvDAT нейроны, которые находились в относительно покоящемся состоянии у нерожавших самок, становились гипервозбудимыми у кормящих агрессивных матерей. У этих самок наблюдалась значительно более высокая частота разрядов и более деполяризованный потенциал покоя.
Для доказательства причинно-следственной связи между активностью PMvDAT нейронов и материнской агрессии ученые использовали методы оптогенетики и генетической абляции. При оптогенетической стимуляции PMvDAT нейронов у кормящих самок наблюдалось немедленное и значительное увеличение продолжительности атак, направленных на «нарушителя». Важно отметить, что фотоактивация этих же нейронов у нерожавших самок не вызывала агрессии, а лишь усиливада исследовательское поведение, что подтверждает зависимость механизма от материнского состояния. Напротив, оптогенетическое ингибирование PMvDAT нейронов у агрессивных самок приводило к уменьшению продолжительности атак и задержке начала эпизодов агрессии. Генетическая абляция PMvDAT нейронов у кормящих самок также резко снижала частоту и продолжительность агрессивного поведения и увеличивала время до первой атаки. Таким образом, активность PMvDAT нейронов является ключевым фактором в материнской агрессии, причем зависит от сомтояния самки.
Затем исследователи выяснили, какие сигналы вызывают переход PMvDAT нейронов в гипервозбудимое состояние. Оказалось, что PMvDAT нейроны чувствительны к материнским гормонам, уровень которых резко возрастает во время лактации, — пролактину и окситоцину. У кормящих мышей уровень пролактина в сыворотке крови был в десять раз выше, чем у нерожавших самок. При этом в PMvDAT нейронах лактирующих самок усиливалось фосфорилирование STAT5, который участвует в сигналинге через рецепторы простагландина. Пролактин увеличивал возбудимость клеток в живых срезах мозга — после обработки в них наблюдалась преждевременная деполяризация, повышение частоты разрядов и удлинение потенциала действия. Окситоцин, хотя он чаще ассоциируется с материнской заботой и социальными связями, также показал способность вызывать обратимую деполяризацию PMvDAT нейронов, аналогичную действию пролактина. Таким образом, пролактин и окситоцин действуют синергично, облегчая в PMvDAT нейронах активацию, характерную для агрессивных особей.
Материнская агрессия и забота о потомстве включаются одновременно после родов. Ученые хотели выяснить, способствует ли этот нейронный контур всем материнским поведенческим программам или же он специализирован. Анализируя, как самка возвращает детенышей в гнездо, они обнаружили, что оптогенетическая активация PMvDAT нейронов сильно нарушает этот процесс даже в отсутствие нарушителя. При этом агрессии по отношению к самим детенышам не наблюдалось. Это говорит о том, что активация PMvDAT нейронов запускает специализированную программу агрессии, подавляя другие формы материнского поведения. При угрозе этот механизм может служить для приоритизации атаки над заботой, позволяя направить ресурсы на активную защиту потомства.
Авторы предполагают, что половой диморфизм в агрессивном поведении обусловлен не принципиально различной организацией нейронных цепей у самцов и самок. Наоборот, существует единая нейронная основа (PMvDAT нейроны), которая в нормальных условиях подавлена у самок, но временно мобилизуется адаптивными гормональными сигналами (пролактином и окситоцином), когда необходимо защищать потомство.
Таким образом, ученые описали механизм, позволяющий животному временно получить доступ к «спящей» поведенческой программе для адаптивных целей. Гормональные перестройки временно обеспечивают гипервозбудимость нейронов, участвующих в агрессивном поведении, и тем самым позволяют самке защищать потомство.
Источник
Stagkourakis, S., et al. //Maternal aggression driven by the transient mobilisation of a dormant hormone-sensitive circuit. Nat Commun 16, 8553 (2025). DOI: 10.1038/s41467-025-64043-4

Вам будет интересно
 130
130
 0
0



 160
0
160
0