
 Меню
Меню





 Все темы
Все темы
Спаривание мышей регулируется сложнее, чем считалось
Исследователи из Португалии и Франции обнаружили, что спинной мозг мышей играет более сложную роль в сексуальном возбуждении, чем предполагалось. Ранее считалось, что он контролирует только рефлекс эякуляции. Однако авторы экспериментально показали, что популяция галанин-положительных (Gal+) нейронов в поясничном отделе спинного мозга (L2/L3) модулирует сексуальное возбуждение и влияет на сам процесс спаривания. Эти нейроны связаны с работой луковично-губчатой мышцы, сокращения которой обеспечивают выброс семени. Gal+-нейроны интегрируют сенсорные сигналы, а их активность отражает общее возбуждение мыши (в том числе в ответ на агрессию). Удаление этих нейронов также нарушало копулятивное поведение самцов.
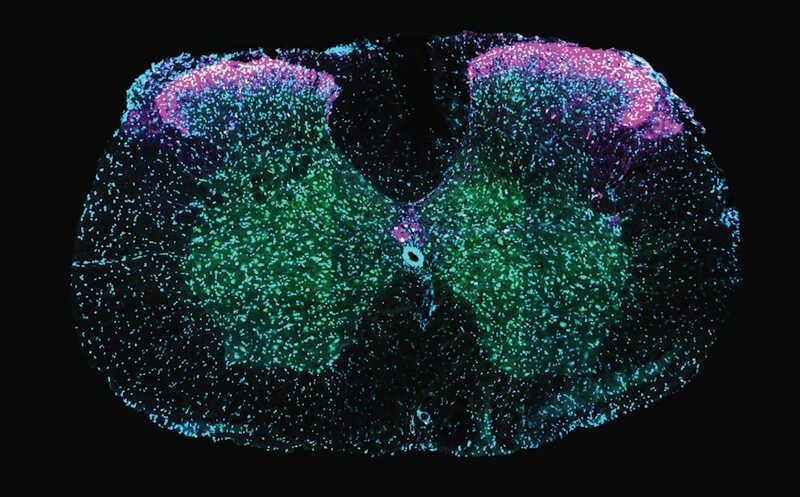
Срез спинного мозга самца мыши после эякуляции: часть окрашенных розовым Gal+-нейронов также несет зеленую метку активации, что подтверждает активность этой популяции нейронов.
Credit:
Neuroethology Lab, Champalimaud Foundation |
подробнее
В классической нейробиологической модели считается, что прекопулятивные действия и копуляция регулируются головным мозгом, в то время как сама эякуляция является двухэтапным рефлексом (эмиссия и выброс семени), полностью контролируемым цепями спинного мозга. Согласно этой концепции, спинной мозг остается под сильным тормозящим влиянием нисходящих сигналов из мозга до тех пор, пока не будет достигнут эякуляторный порог. Однако то, как именно копулятивные последовательности организуются и как мозг «сообщает» спинному мозгу о достижении этого порога, оставалось не до конца ясным.
Чтобы разобраться в механизмах, стоящих за эякуляцией, исследователи картировали у самцов мыши спинномозговую цепь, которая контролирует бульбоспонгиозную, или луковично-губчатую мышцу (bulbospongiosus muscle), необходимую для выброса спермы. Ученые вводили в бульбоспонгиозную мышцу флуоресцентную ретроградную метку, чтобы идентифицировать мотонейроны, ответственные за сокращение этой мышцы. Они располагались преимущественно в сегментах L6 и S1 пояснично-крестцового отдела спинного мозга.
Затем, применив трассирование с помощью вируса псевдобешенства, авторы идентифицировали пресинаптические партнеры этих мотонейронов. Обнаруженный плотный кластер нейронов, экспрессирующих галанин (Gal+), располагался вокруг центрального канала в сегментах спинного мозга L2/L3. Используя Cre-рекомбинацию для направленного внесения меток и метод пэтч-кламп, исследователи доказали наличие функциональной моносинаптической связи между Gal+ нейронами и мотонейронами луковично-губчатой мышцы. Передача сигнала осуществляется через глутаматергические синапсы. Кроме того, Gal+ нейроны анатомически связаны с симпатическими и парасимпатическими центрами, что предполагает их роль не только в выбросе спермы, но и в эрекции.
Чтобы понять, как спинномозговая цепь, контролирующая выброс спермы, реагирует на внешние стимулы, исследователи провели на самцах мыши опыты с сенсорной и оптогенетической стимуляцией. Во-первых, они исследовали, получают ли мотонейроны луковично-губчатой мышцы и Gal+ нейроны сенсорный сигнал от пениса. Используя стимуляцию струей воздуха, ученые обнаружили, что стимуляция пениса вызывает значительно большие локальные потенциалы в этих спинномозговых областях, чем стимуляция лапы. Эти результаты стали функциональным доказательством того, что обе популяции нейронов получают сенсорные сигналы из области гениталий.
Во-вторых, исследователи стимулировали Gal+ нейроны (электрически или оптогенетически), чтобы проверить, может ли это вызвать активность луковично-губчатой мышцы. Выраженная активность мышцы наблюдалась только в случае, когда была нарушена связь спинного мозга с головным. У интактных мышей стимуляция приводила лишь к минимальному ответу. Это подтверждает, что Gal+ нейроны и их сенсорный вход находятся под сильным тоническим нисходящим торможением со стороны мозга. Предполагается, что это торможение — механизм, предотвращающий непроизвольную эякуляцию до достижения необходимого порога возбуждения.
Интересно, что ответ луковично-губчатой мышцы на электрическую стимуляцию Gal+ нейронов зависел от внутреннего состояния самца. У самцов, которые перед экспериментом начали спариваться с самкой, однако интромиссия была прервана до эякуляции, луковично-губчатая мышца сильнее отвечала на стимуляцию, чем у спокойных или недавно эякулировавших самцов. Кроме того, многократная стимуляция Gal+ нейронов вызывала уменьшение амплитуды и длительности ответа, что не наблюдалось при прямой стимуляции мотонейронов мышцы. Это указывает на то, что свойства нейронных цепей Gal+ модулируются копуляцией и эякуляцией.
Для дальнейшего анализа роли Gal+ нейронов в возбуждении, ученые измерили активацию этих клеток во время спаривания. Измерение проводили по экспрессии гена cFos, маркера нейронной активности. Как и ожидалось, в группе самцов, достигших эякуляции, наблюдалось наибольшее количество активных Gal+ нейронов. Однако Gal+ нейроны также повышали активность при половом возбуждении и даже при взаимодействии с другим агрессивным самцом — по-видимому, их активность связана с общим уровнем внутреннего возбуждения, а не только со специфическим сексуальным взаимодействием.
Чтобы окончательно определить функцию этой популяции нейронов, исследователи провели эксперимент по их генетической абляции. Самцам мышей вводили вирусный вектор, кодирующий рецептор дифтерийного токсина, в спинные сегменты L2/L3. После введения дифтерийного токсина нейроны Gal+ погибали. Абляция Gal+ нейронов значительно нарушила последовательность копулятивного поведения —у самцов значительно увеличилось время до эякуляции, однако время, необходимое для начала акта спаривания, не изменилось. Также возрастало число неглубоких тазовых движений, когда самец ищет влагалище самки, а также увеличилась продолжительность этого поиска.
Важно отметить, что, когда эякуляторный порог был достигнут, продолжительность самой эякуляции оставалась нормальной. Эти результаты, указывающие на сбой в организации копулятивной последовательности, резко контрастируют с данными, полученными на крысах, где абляция аналогичной группы нейронов влияла только на рефлекс эякуляции, оставляя копуляцию нетронутой.
Таким образом, спинной мозг играет более сложную и центральную роль в половом поведении мышей, чем считалось ранее. Спинальная цепь Gal+ нейронов не является простым «переключателем» для рефлекса эякуляции, а активно участвует в интеграции сенсорных сигналов и отражает внутреннее состояние возбуждения животного.
Поскольку активность этих нейронов зависит от поведенческого состояния — самец спокоен, возбужден или после эякуляции, — это позволяет предположить, что спинальная цепь также может быть вовлечена в контроль рефрактерного периода.
Исследование подтверждает гипотезу о постоянном «диалоге» между мозгом и спинным мозгом во время сексуального возбуждения и копуляции. В отличие от крыс, чья спинальная цепь может работать как истинная рефлекторная дуга, способная вызвать эякуляцию только за счет сенсорного ввода при удалении нисходящего торможения, у мышей необходим постоянный обмен информацией с вышестоящими мозговыми центрами для успешного спаривания.
Диета контролирует половое поведение дрозофилы
Мыши в спокойной обстановке предпочитают контактировать с самками, а при угрозе — с самцами
Источник
Lenschow, C., et al. A galanin-positive population of lumbar spinal cord neurons modulates sexual arousal and copulatory behavior in male mice. // Nat Commun 16, 8282 (2025). DOI: 10.1038/s41467-025-63877-2
Вам будет интересно

 125
125
 0
0


 157
0
157
0








