Меню
Меню





 Все темы
Все темы
Антиген заставляет CAR T-клетку делиться несимметрично
Для эффективной CAR T-клеточной терапии важны долгоживущие CAR T-клетки памяти. Авторы статьи в Nature установили, что они дифференцируются из введенных пациенту CAR T-клеток одновременно с эффекторными клетками. При активации антигеном CAR T-клетка делится асимметрично, и одна из дочерних клеток становится короткоживущей эффекторной клеткой, а вторая — клеткой памяти.
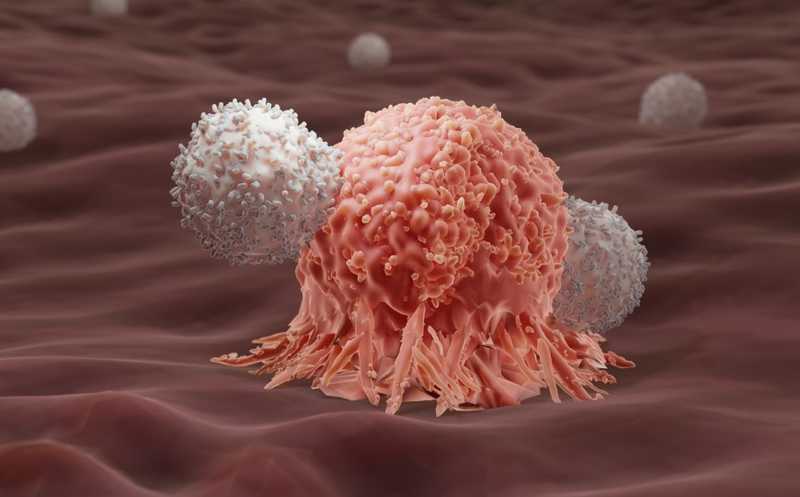
Успехи CAR T-клеточной терапии связаны с длительной персистенцией терапевтических Т-клеток в организме пациента. Она объясняется образованием CAR T-клеток памяти — долгоживущих лимфоцитов, которые обеспечивают стабильный иммунологический надзор и тем самым предотвращают рецидивы рака. Однако механизмы, определяющие судьбу дифференцировки делящихся CAR-T клеток, остаются неясными. Один из возможных путей формирования памяти после активации CD8+ T-клеток — это их асимметричное деление, уже выявленное в мышиной модели. Авторы статьи в Nature установили, что оно индуцируется мишенью, с которой контактирует CAR T-клетка. При этом CAR, распознавший мишень, остается на поверхности одной из дочерних клеток, и та дифференцируется в короткоживущую эффекторную Т-клетку, а вторая приобретает фенотип памяти.
Для отслеживания судьбы дифференцирующихся клеток исследователи воспользовались методом LIPSTIC (labelling immune partnerships by SorTagging intercellular contacts) — бактериальный фермент сортаза А в нем обеспечивает перенос меченного субстрата между CD40L и CD40 на взаимодействующих иммунных клеток. При взаимодействии CAR-T с клеткой-мишенью происходит обогащение метки LIPSTIC в иммунном синапсе, после чего она специфически переносится с мишени на CAR-T. Проточная цитометрия дочерних CAR T-клеток после первого деления выявила равные доли LIPSTIC+ и LIPSTIC– клеток, что свидетельствует об асимметричном распределении метки. Визуализация клеток во время митоза подтвердила, что метка сохраняется исключительно на одной дочерней клетки, тогда как вторая ее не получает.
Асимметрия затрагивала не только распределение метки, но и фенотип клеток. LIPSTIC+ дочерние клетки были метаболически активнее и переходили к гликолизу, что способствовало увеличению их размера и ускоряло пролиферацию. В отличие от них, LIPSTIC– дочерние клетки полагались преимущественно на окислительное фосфорилирование. Первые из них были обозначены как проксимальные, вторые — как дистальные дочерние CAR-T.
Авторы работы проверили, свойственно ли асимметричное деление различным подтипам CAR T-клеток. Опыты in vitro показали, что асимметрично делятся как CD4+, так и CD8+ CAR-T. При сравнении эффекторных и наивных Т-клеток выраженная асимметрия обнаружилась среди дочерних клеток последних, хотя и у потомков эффекторных CAR-T наблюдалась подобная тенденция. Способность к асимметричному делению не только наблюдалась при первом митозе, но и сохранялась у последующих поколений.
Функциональное значение такой асимметрии деления CAR T-клеток исследователи проанализировали in vivo. Они вводили отсортированные проксимальные или дистальные дочерние клетки NSG-мышам и выяснили, что через 30 дней после инъекции Т-клеток в периферической крови было больше у тех животных, которые получили дистальные дочерние CAR-T. Через 35 дней после инъекции мышам вводили также клетки линии Nalm6 в качестве мишеней. Распространение лейкоза в такой модели были способны контролировать только дистальные дочерние клетки. Кроме того, в этом случае в селезенке присутствовало больше Т-лимфоцитов, чем при введении проксимальных дочерних CAR-T, и у них сохранялся фенотип наивных клеток — это говорит о самообновлении таких CAR-T in vivo, тогда как проксимальные дочерние клетки оказались короткоживущими.
Анализ цитотоксичности in vitro показал, что проксимальные дочерние клетки активно убивают опухолевые при очень низком соотношении эффектор:мишень. Неожиданно дистальные CAR T-клетки тоже продемонстрировали мощную цитотоксическую активность.
Опыты на мышиной модели лейкоза подтвердили цитотоксичность обоих типов дочерних клеток. Первоначальная опухолевая нагрузка снижалась и в том, и в другом случае, однако у мышей, получавших проксимальные дочерние CAR T-клетки, контроль над развитием лейкоза постепенно терялся.
Далее исследователи задались вопросом, какие механизмы определяют функциональное расхождение дочерних клеток, формирующихся при несимметричном делении активированного CAR T-лимфоцита. Проанализировав экспрессию поверхностных белков, транскрипционную активность и клональность T-клеточного рецептора в единичных клетках, они пришли к выводу, что во время первого клеточного деления закладывается ассимметрия протеома поверхности. Так, на поверхности дистальных дочерних клеток присутствовало больше CD45RA, на поверхности проксимальных — CD25. Кроме того, дистальные CAR-T активнее экспрессировали CD5, ингибирующий активность и пролиферацию T-клеток.
Анализ обогащения наборов генов по данным транскриптомики выявил в дистальных дочерних клетках ассоциированные с наивным фенотипом сигнатуры; экспрессия эффекторных генов у них была подавлена. В проксимальных дочерних клетках активнее экспрессировались гены-мишени MYC и MTORC1, а также компоненты гликолиза. Такая транскрипционная асимметрия, как выяснилось, обеспечивается двумя механизмами — неравномерным распределением РНК между дочерними клетками и регуляцией генов после деления.
Авторы работы сравнили активность различных регуляторов между проксимальными и дистальными дочерними CAR T-клетками. В проксимальных они обнаружили активацию путей, связанных с апоптозом, пролиферацией и дифференцировкой эффекторных клеток. Напротив, в дистальных дочерних клетках повышалась активность транскрипционных факторов, связанных с самообновлением и подавлением пролиферации, таких как KLF2, IRF1, STAT1/2, REST и IKZF1. Последний представляет собой транскрипционных фактор, который подавляет экспрессию интерлейкина-2 (IL-2) и его рецептора (IL2RA). Нокаут IKZF1 в CAR T-клетках привел к тому, что дистальные дочерние клетки приобретали фенотип проксимальных. Аналогичный эффект наблюдался in vivo — в организме модельных мышей CAR-T, лишенные этого гена, теряли способность к эффективному и долговременному сдерживанию опухоли.
Данная работа демонстрирует, что активация CAR T-клетки мишенью стимулирует асимметричное деление, благодаря которому формируются две популяции дочерних клеток с разными функциями — дистальные, работающие как Т-клетки памяти, и проксимальные — эффекторные. Важным транскрипционным фактором, определяющим судьбу дистальных дочерних CAR T-клеток и способствующим их цитотоксичности и персистенции, служит IKZF1.
CAR-T клетки стали эффективнее с мутациями, найденными в лимфоме
Источник

Вам будет интересно



 164
164
 0
0