Меню
Меню





 Все темы
Все темы
Нобелевская неделя 2025. Симон Сакагути о том, как были найдены регуляторные Т-клетки
«Потенциально аутореактивные Т-лимфоциты присутствуют у каждого здорового индивида, но постоянно подавляются регуляторными Т-клетками (Treg). Граница между “своим” и “не-своим” четко не зафиксирована. Она может смещаться в зависимости от числа Treg. Аутоиммунные заболевания можно лечить, противоопухолевый иммунитет — вызывать, а трансплантационную толерантность — устанавливать, контролируя количество и функцию Treg». — Лекция лауреата Нобелевской премии 2025 года по физиологии и медицине Симона Сакагути. Стокгольм, 07.12.2025.

Скриншот канала Нобелевского комитета
Что защищает нас от «ужаса аутоинтоксикации»
Свое выступление о роли регуляторных Т‑клеток в иммунологической толерантности Симон Сакагути начал с напоминания о том, что основная задача иммунной системы — защищать организм от вторгающихся микробов. Она обладает уникальными свойствами: Т- и B-клетки, с высокой специфичностью реагирующие на чужеродные антигены, иммунологическая память, мощные эффекторные механизмы, такие как выработка антител и цитотоксическая активность Т‑клеток. Однако эти «инструменты», как правило, не реагируют на собственные антигены организма.
В начале XX века Пауль Эрлих ввел термин horror autotoxicus («ужас аутоинтоксикации»). Он предположил, что должны существовать определенные регуляторные механизмы, которые защищают нас от развития аутоиммунной реакции, — ведь в норме она не происходит.
С тех пор было предложено несколько таких механизмов. Первый — делеция клонов: незрелые Т-или В-клетки, которые распознают собственные антигены, подвергаются апоптозу. Вторая идея — инактивация через отсутствие подтверждающего сигнала. Для полной активации Т‑клетки требуется не только распознавание антигена через Т-клеточный рецептор, но и костимуляция; при отсутствии этого второго сигнала Т-клетки остаются неактивными и происходит их анергия. И третий механизм — супрессия. Даже у здоровых людей могут присутствовать аутореактивные лимфоциты, но их активация и пролиферация контролируются другой субпопуляцией Т-клеток — тех самых, которые сейчас называются регуляторными Т-клетками (Treg).
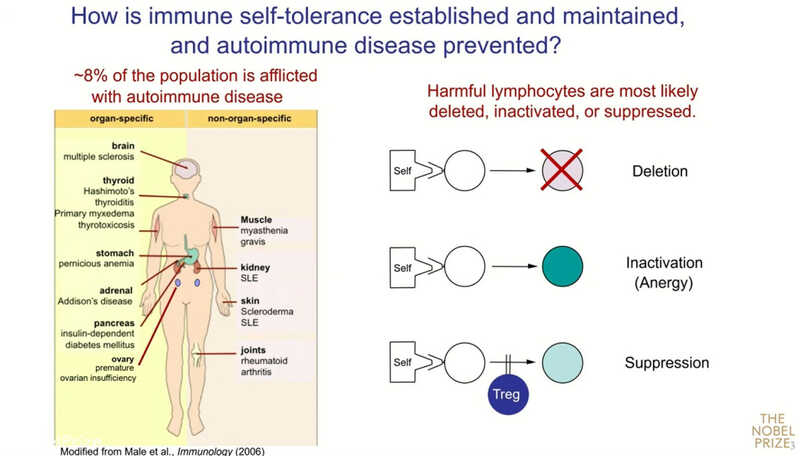
Тяжелые и широко распространенные аутоиммунные заболевания — рассеянный склероз, аутоиммунный гастрит, тиреоидит Хашимото, поражение поджелудочной железы (инсулинзависимый сахарный диабет) или ревматоидный артрит. Аутоиммунные процессы могут затрагивать репродуктивную систему и вызывать бесплодие как у женщин, так и у мужчин. Примерно 8% населения Земли страдают от того или иного аутоиммунного заболевания.
Экспериментальные основания открытия Treg
Еще в 1969 году патологи из Японии Ясуаки Нисидзука и Теруо Сакакура обнаружили, что если удалить тимус у мышат примерно на третий день после рождения, то мыши страдают от аутоиммунных заболеваний. В 1980-е годы эти исследования продолжил Симон Сакагути с коллегами.
Например, у трети мышей линии А после тимэктомии, выполненной на третий день жизни, развивается аутоиммунный гастрит, практически с той же частотой — поражение яичников у самок. Интересно, что если провести тимэктомию в течение первых 24 часов после рождения или позднее — на седьмой день жизни, то эффекта не наблюдается. У мышей линии В выполненная на 3-й день тимэктомия приводила к тем или иным аутоиммунным заболеваниям уже примерно в 90% случаев. Но у мышей линии С удаление тимуса на том же сроке не вызывало аутоиммунных нарушений вовсе.
Таким образом, тимэктомия именно на третий день после рождения является ключевым моментом. Кроме того, в зависимости от генетического фона развитие аутоимунных заболеваний происходит с разной частотой. Как тимэктомия в раннем неонатальном периоде может вызвать аутоиммунные заболевания и почему критичен трехдневный срок?

«Что мы обнаружили: если после неонатальной тимэктомии инокулировать таким мышам нормальные Т-клетки от взрослых мышей, особенно CD4⁺ Т-клетки, то развитие аутоиммунных заболеваний можно предотвратить, — рассказывает нобелевский лауреат. —Наша гипотеза заключалась в том, что поскольку при тимэктомии на третий день после рождения развивается аутоиммунитет, это означает, что, возможно, до этого тимус уже успел сгенерировать аутоиммунные Т-клетки. Примерно к трем суткам он начинает вырабатывать другую, особую популяцию — клетки, супрессирующие аутоиммунные реакции. Сегодня мы называем их регуляторными Т‑клетками.
Таким образом, если провести тимэктомию в первый день, не будет никаких Т-клеток и аутоиммунитета. Если же тимэктомию провести на седьмой день, уже появляется достаточное количество регуляторных Т‑клеток на периферии, и аутоиммунные заболевания также не возникают».
Молекулярные маркеры Treg
Доктор Сакагути с коллегами поставили такие вопросы: как можно разделить регуляторные Т-клетки и аутоиммунные Т-клетки у нормальных взрослых мышей? И если удалить Treg у нормальных взрослых мышей, получится индуцировать аутоиммунное заболевание или нет?
У нормальных мышей в селезенке содержится огромное количество разнообразных Т-клеток. «Мы начали дробить эту популяцию Т-клеток селезенки, используя как маркеры молекулы, экспрессируемые на поверхности клеток. Как убрать их? С помощью антител и комплемента», — объясняет Симон Сакагути. Антитело связывается с определенной молекулой на поверхности клетки, комплемент активируется — и клетки лизируются. После такой обработки оставшиеся клетки переносят мышам, у которых Т‑клетки отсутствуют вообще. И если у таких мышей развивается аутоиммунное заболевание, скорее всего, это говорит о том, что удаленная популяция обладала супрессивной активностью и подавляла развитие аутоиммунитета.
Первым таким успешным маркером стала молекула CD5. Пятью годами позже, в 1991 году, Фиона Паури и Дон Мэйсон из Оксфордского университета (Великобритания) показали, что маркером клеток с иммунорегуляторными свойствами может служить сниженное количество изоформы CD45 — CD45RB. Но тогда перед исследователями возник следующий вопрос: можно ли найти более специфичный маркер для этой популяции клеток? В итоге остановились на CD25.
На слайде лимфоциты показаны кружочками, а CD4⁺ CD25⁺-клетки перечеркнуты — именно эту популяцию удаляли с помощью антител к CD25 и комплемента. Если оставшиеся клетки (CD25⁻) ввести мышам, лишенным Т-клеток, то у них начнутся аутоиммунные заболевания.
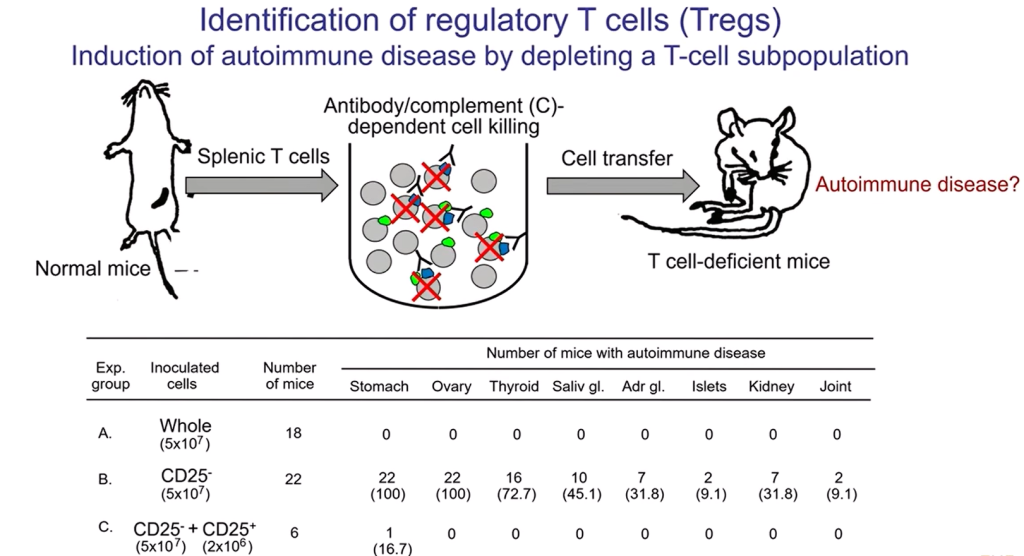
Если переносить мышам все Т-клетки без отбора, ничего не происходит. Если же переносить только CD25⁻ клетки, то у 100% животных развиваются аутоиммунный гастрит, поражение яичников и другие патологии.
«Мы можем продемонстрировать их супрессивную активность in vitro. В селезенке около 10% всех CD4⁺ Т‑клеток положительны по CD25, а в тимусе их от 5 до 10%. Если удалить CD25⁺ клетки в селезенке, то CD25⁻ клетки начинают активно размножаться в ответ на собственные антигены. А если в такую культуру добавить обратно CD25⁺ клетки, то пролиферация CD25⁻ клеток сильно подавляется, причем эффект является дозозависимым. Они подавляют даже пролиферацию CD8⁺ Т-клеток. Также и в тимусе CD25⁺ клетки способны подавлять пролиферацию других лимфоцитов», — рассказывает Симон Сакагути.
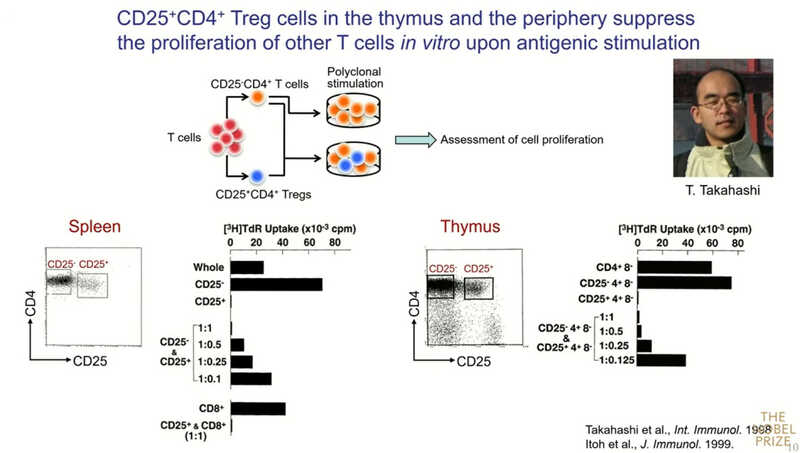
Эти данные позволили предположить, что Т-клетки, продуцируемые тимусом, функционально разнородны, и среди них существует отдельная супрессивно-компетентная популяция. Они дифференцируются в зрелую популяцию в тимусе, им требуется больше времени для созревания, а выход в периферию задерживаются. Именно поэтому тимэктомия на третий день жизни мышонка вызывает аутоиммунные заболевания: она удаляет только регуляторные Т-клетки.
«Т-регуляторные клетки тимуса связаны как с центральной, так и с периферической толерантностью. В тимусе большинство высокоаутореактивных клеток удаляются через клональную негативную селекцию, но некоторые из них, с промежуточным сродством к собственным антигенам, могут каким-то образом дифференцироваться в Treg, — объясняет Симон Сакагути. — Низкоафинные к собственным антигенам клетки могут избежать негативной селекции и мигрировать на периферию. Такие клетки, если мы удалим Treg, способны активироваться и вызвать аутоиммунное заболевание».
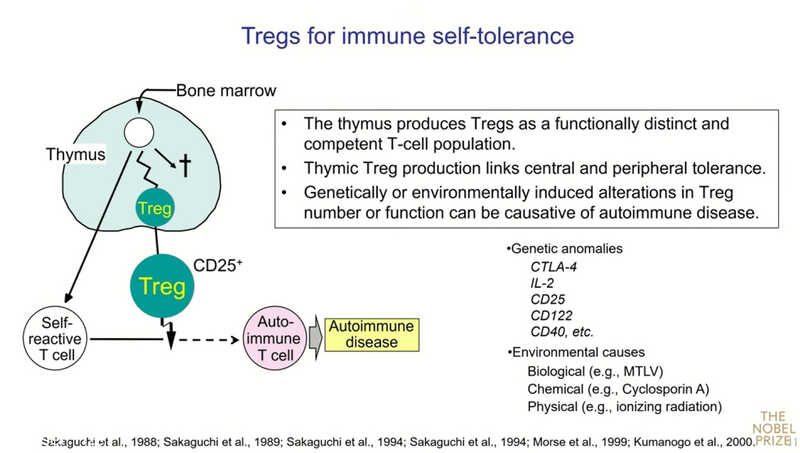
Аутоиммунные заболевания у людей и мышей и ген Foxp3
Следующий вопрос: работает ли обнаруженный механизм у людей? Важные данные были получены Мэри Брункоу и Фредом Рамсделлом в 2001 году. Они показали, что мутации в гене, который они назвали Foxp3, приводят к редкому заболеванию у человека — IPEX-синдрому (Immune dysregulation, Polyendocrinopathy, Enteropathy, X-linked syndrome). Это моногенное заболевание сопровождается аутоиммунным поражением множества органов. У детей развиваются сахарный диабет 1 типа, тиреоидит, тяжелое воспалительное заболевание кишечника, а после введения прикорма начинаются аллергии. Сходная картина заболевания была у мышей линии scurfy, с изучения которых началась работа Брункоу и Рамсделла.
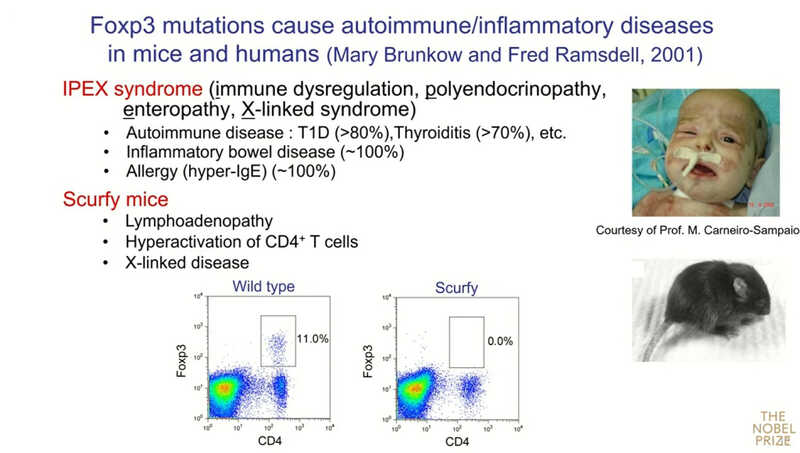
«Нас заинтересовал этот ген, потому что клиническая картина IPEX была удивительно похожа на ту, что мы наблюдали у мышей после удаления регуляторных Т-клеток. Поэтому мы получили антитело к Foxp3 и нашли, что в норме у мышей примерно 10% CD4⁺ Т-клеток экспрессируют Foxp3, но у больных мышей не было таких клеток. Сейчас мы можем окрашивать Т-регуляторные клетки по маркерам Foxp3 и CD25». Подавляющее большинство клеток, экспрессирующих Foxp3, одновременно положительны по CD25 — как в селезенке, так и в тимусе.
Что это означает? Во-первых, потенциально аутореактивные клетки присутствуют у каждого здорового человека, но их активность подавляется Treg. Во-вторых, граница между «своим» и «не-своим» четко не определена. Она может перемещаться в зависимости от численности и функции регуляторных Т‑клеток. Можно сместить эту границу, усилив или ослабив Treg. Следовательно, регулируя количество и функции Treg, мы можем лечить аутоиммунные заболевания, вызывать иммунитет к опухолям или поддерживать толерантность к трансплантату.
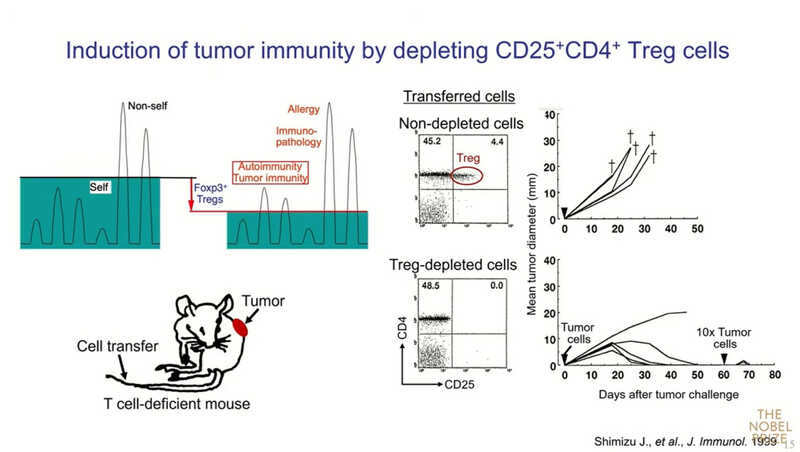
Если мышам ввести опухолевые клетки, то опухоль бесконтрольно растет, и все они погибают. Но когда из организма мышей предварительно удаляли Treg, перенесенная опухоль сначала росла, а потом отторгалась. Если же ввести животному опухолевые клетки в 10‑кратной дозе, то опухоль отторгается мгновенно.
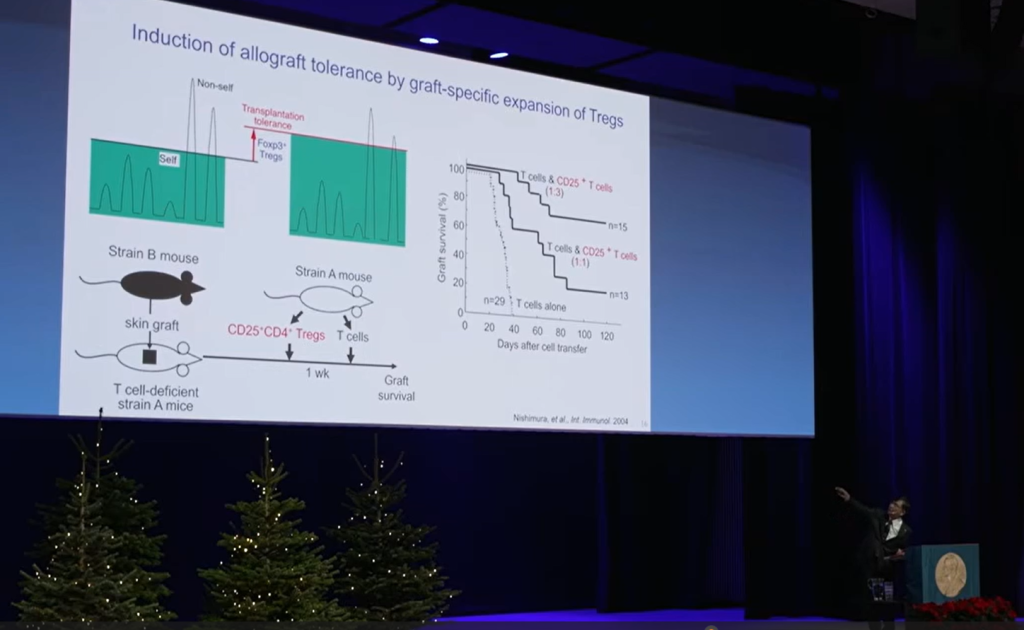
А что с трансплантацией? Мышь с дефицитом Т-клеток может принять кожный трансплантат от другого штамма мышей на 100%. Если ввести этой мыши нормальные Т-клетки, то отторжение происходит в течение месяца. Но если за неделю до этого ввести ей регуляторные Т-клетки, то они мигрируют в лимфоидные органы и подавляют отторжение. У 30% мышей трансплантат приживается навсегда, а при увеличении дозы Treg более 70% становятся постоянно толерантными без назначения иммуносупрессивных препаратов.
Молекулярные основы работы Т-регуляторных клеток
«Foxp3 контролирует сотни генов: или активирует, или подавляет. Например, CD25 и CTLA-4 активируются, а IL-2 и IFN-γ— подавляются. Вопрос: какие именно гены среди контролируемых Foxp3 важны для супрессивной активности? При дефиците какого гена у мышей развивается тяжелое аутоиммунное заболевание, какое мы видим при мутации Foxp3? Их не так много. Одна группа — IL-2 и его рецептор. Другая— CTLA-4».
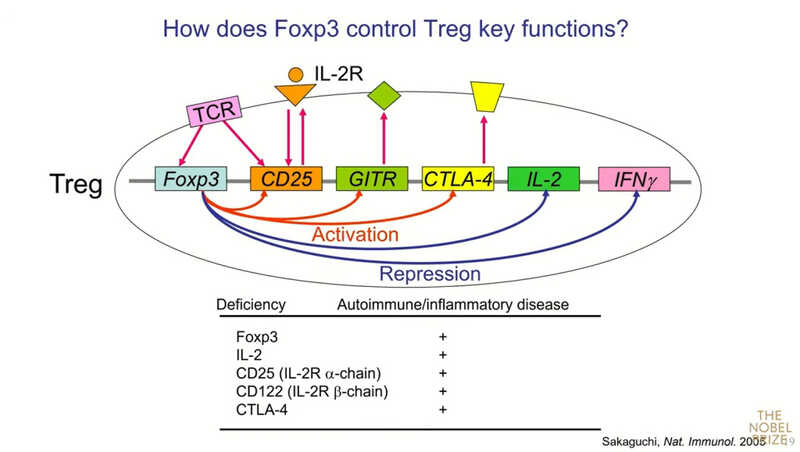
Ключевая особенность Treg — то, что они конститутивно экспрессируют высокоафинный рецептор к IL-2 (это и есть CD25). Его экспрессия начинается, когда клетки еще находятся в тимусе. Foxp3 подавляет продукцию самого IL-2. Когда активируются обычные Т-лимфоциты, то они продуцируют IL-2 и рецептор к нему. Регуляторные Т‑клетки экспрессируют только рецептор к IL-2, но не сам IL-2. Их созревание и выживание полностью зависит от внешнего IL-2. Поэтому, если ввести нейтрализующее антитело к IL-2, то почти немедленно можно увидеть резкое сокращение числа Treg как в селезенке, так и в тимусе, а затем тяжелое аутоиммунное заболевание.
Кайса Винг из Швеции создала линию нокаутных мышей с дефицитом Treg-специфичного CTLA-4. Foxp3-положительные клетки этих мышей не экспрессировали CTLA-4, и у них спонтанно развивались очень тяжелые аутоимунные заболевания, например, миокардит. Иными словами, дефицит CTLA-4 у регуляторных Т-клеток вызывает те же патологии, что и удаление самих Treg — это важная молекула для их функции.
Нобелевский лауреат коротко описал актуальные представления о том, какие молекулы экспрессируются регуляторными T-клетками и как они участвуют в иммуносупрессии. CTLA-4 взаимодействует с CD80 и CD86 — так называемыми костимуляторными молекулами на поверхности антигенпрезентирующих клеток (АПК). У эффекторной клетки распознавание антигена наряду с костимуляцией вызывает активацию и деление. Treg конкурируют с эффекторными клетками за CD80/CD86 с помощью CTLA-4 и снижают экспрессию, буквально забирая их с поверхности АПК путем транс-эндоцитоза. То есть, во-первых, Т-регуляторные клетки напрямую подавляют костимуляторный сигнал.
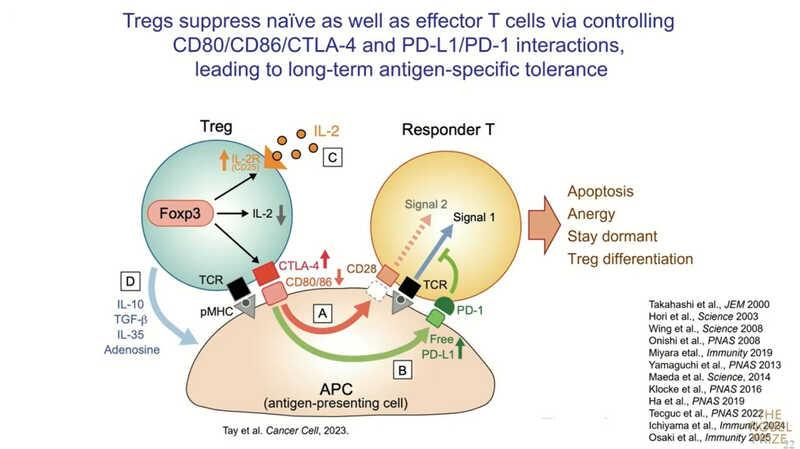
Во-вторых, происходит взаимодействие с PD-L1. Этот важный регулятор также является хорошей мишенью для иммунотерапии. В норме он ассоциирован с CD80. Когда количество CD80 уменьшается, концентрация свободного PD-L1 возрастает, и он взаимодействует с PD-1-позитивными Т-клетками. Таким образом Treg могут подавлять активированные PD-1-позитивные Т-клетки.
В-третьих, при дальнейшей дифференцировке Т-регуляторные клетки могут продуцировать иммуносупрессивные цитокины, основные мишени которых — АПК.
И, наконец, воздействие на IL-2: так как Treg конститутивно экспрессируют рецептор IL-2 (CD25), но сам IL-2 не продуцируют, они активно поглощают его из микроокружения, лишая эффекторные Т‑клетки этого важного фактора роста.
«Tregопатии» и Treg-лекарства
При моногенных заболеваниях мутации в генах, важных для регуляторных Т-клеток, таких как Foxp3, Cd25 и Ctla4, нарушают их развитие и функцию. Такие состояния можно назвать «Tregопатиями», и они сходны по клинической картине с синдромом IPEX.
При полигенных заболеваниях в энхансерных регионах генов Foxp3, Cd25 и Ctla4 обнаруживаются однонуклеотидные полиморфизмы (SNP). Эти SNP способствуют генетической предрасположенности к распространенным аутоиммунным заболеваниям, таким как сахарный диабет 1 типа и ревматоидный артрит.
Так что с терапией аутоиммунных заболеваний? Во-первых, можно увеличить количество Treg in vivo. Поскольку они экспрессируют рецептор с высоким сродством к IL-2, достаточно ввести низкие дозы рекомбинантного IL-2, и этого хватит для селективного увеличения популяции Treg, но не эффекторных клеток. Так можно усилить иммунологическую толерантность. Второй подход — получить Treg из крови пациента, увеличить ex vivo, и вернуть обратно.
«Третий подход — тот, над которым мы сейчас работаем, — превратить антигенспецифические Т‑клетки в регуляторные in vivo или in vitro, — говорит Симон Сакагути. — Для того, чтобы модифицировать эффекторные Т-клетки, нужно внести в них одновременно Foxp3 и некоторые эпигенетические модификации, характерные для регуляторных клеток. Так можно получить очень стабильные Treg, и введя их обратно пациенту, подавить развитие болезни». Иначе говоря, те самые клетки, которые вызывают аутоиммунное заболевание, превращаются в терапевтические клетки. (Подробнее на PCR.NEWS.)
Сейчас эти методы выходят в клинику. Увеличивая количество Treg и усиливая их функцию, уже возможно лечить аутоиммунные заболевания, аллергию, иммунопатологические заболевания, трансплантировать органы без отторжения и поддерживать толерантность матери к плоду, чтобы предотвратить самопроизвольный аборт на ранних сроках беременности. С другой стороны, снижая количество Treg и ослабляя их супрессивную функцию, можно усилить иммунный ответ к опухолям или при хронических инфекциях.
Завершая выступление, Симон Сакагути поблагодарил тех, кто внес вклад в эту работу: свою жену и коллегу Норико Сакагути, своих учителей Ясуаки Нисидзуку и Теруо Сакакуру, которые познакомили его с подходом неонатальной тимэктомии, Тосияки Такахаси, который принимал участие в исследованиях по разделению популяций Т-клеток с помощью антител, а также других коллег, студентов, коллабораторов и организации, которые финансировали их работу.



 0
0