Меню
Меню





 Все темы
Все темы
Мария Ломакина: «Миграционно пластичные опухолевые клетки могут всё»
На очередной лекции научно-просветительского проекта «Метафаза», который организует PCR.NEWS совместно с парком «Зарядье», о клеточной миграции рассказала Мария Ломакина. Она объяснила, какие способы передвижения доступны некоторым клеткам нашего организма и каким образом те или иные структуры цитоскелета позволяют им выпускать ламеллиподии и блебы или «шагать» от одной точки прикрепления к другой. Клетки опухоли могут приобретать способность к перемещению, за счет чего они преодолевают «препятствия» в организме при метастазировании и инвазии.

Мария Ломакина, кандидат биологических наук, старший научный сотрудник лаборатории механизмов канцерогенеза НИИ канцерогенеза ФГБУ «НМИЦ онкологии Н.Н. Блохина», выступила с лекцией «Клеточная миграция и ее роль в патогенезе опухолей».
Разновидности клеточной миграции
Одно из свойств живого организма — способность к движению, и отдельные клетки внутри организма также могут передвигаться. Клеточная миграция необходима для образования тканей при эмбриогенезе, для поддержания их гомеостаза, заживления ран и обеспечения иммунного ответа. Однако клеточная миграция лежит и в основе патологических процессов, например, развития опухолей, их инвазии и метастазирования.
Клетки способны перемещаться как коллективно, так и поодиночке, причем выделяют мезенхимальный и амебоидный способы индивидуальной миграции. Коллективная миграция, при которой клетки перемещаются «островками», характерна для процессов, связанных с развитием тканей и заживлением ран. По отдельности перемещаются в основном фибробласты, иммунные клетки и клетки крови.
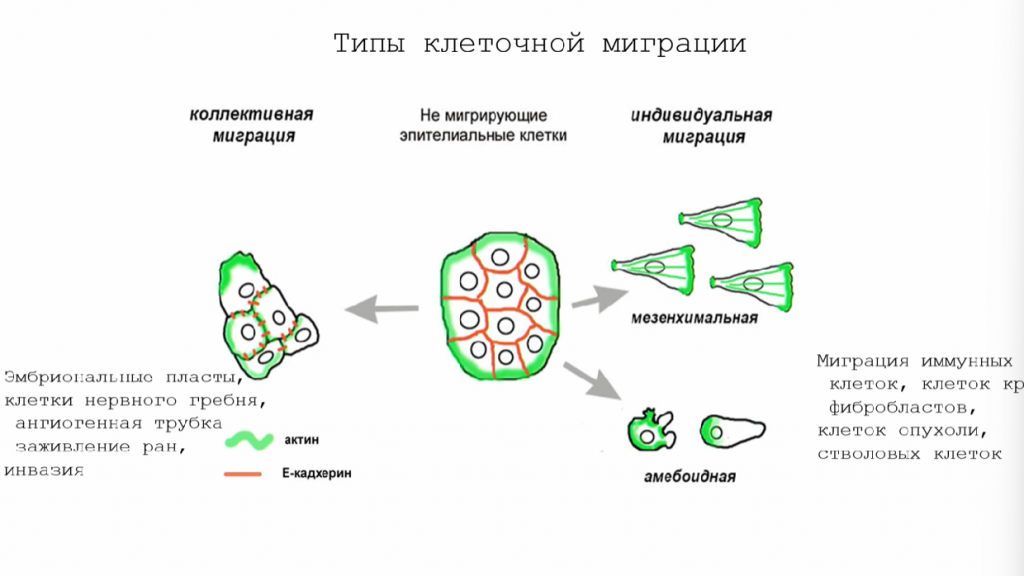
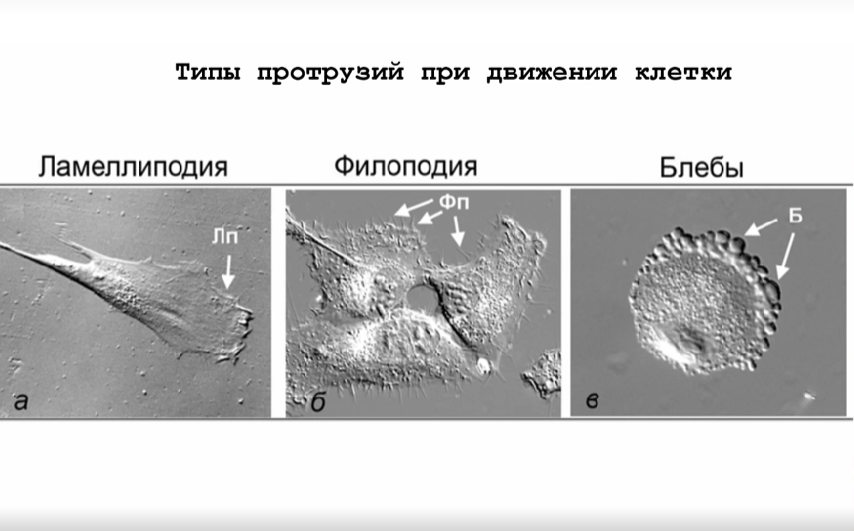
Мигрирующие клетки изменяют свою морфологию так, чтобы у них появлялись выросты разного вида. Клетки, перемещающиеся мезенхимальным способом, образуют ламеллиподии и филоподии, а клетки, для которых характерен амебоидный тип передвижения, — блебы, похожие на пузырьки.
Мезенхимальное движение клеток
Мезенхимальная миграция изучена лучше всего. Большой вклад в ее исследование внес британский биолог-эмбриолог Майкл Аберкромби (1912–1979). Этот ученый впервые описал этапы мезенхимальной миграции: поляризация клетки в ответ на сигнал (появление различий между «передним» и «задним» концом по направлению движения); образование псевдоподий на переднем крае клетки; закрепление псевдоподий и формирование контактов клетки с субстратом; сокращение и подтягивание тела клетки вперед.
Все эти процессы на молекулярном уровне обеспечиваются цитоскелетом, который состоит из актиновых микрофиламентов, промежуточных филаментов и микротрубочек. Из всех этих структур для движения наиболее важны актиновые микрофиламенты; докладчица сравнила их с мышцами нашего организма. Актин — белок, способный полимеризоваться в полярные волокна, которые растут с (+)-конца и разбираются с (–)-конца. Именно из его филаментов, лежащих параллельно друг другу или образующих ветвистые структуры, формируются ламеллиподии и филоподии.
В ходе патологического процесса клетки могут образовывать похожие на филоподии структуры. Например, инвазирующие опухолевые клетки формируют инвадоподии (аналог у некоторых здоровых клеток — подосомы). С инвадоподиями взаимодействуют матриксные металлопротеиназы, помогающие клетке «продираться» сквозь субстрат.
Еще одна интересная структура в цитоскелете клетки — это стресс-фибриллы. Они представляют собой крупные пучки из актина и немышечного миозина 2 типа, способные к сокращению. Стресс-фибриллы, обеспечивающие перенос тела клетки при движении, крепятся к фокальным контактам клетки с субстратом, которые имеют в своем составе большое количество белков (интегрины, паксилин, таллин, зиксин и др). Фокальные контакты обеспечивают взаимодействие клетки с ее микроокружением, осуществляют механосенсорные функции, а также в них происходит выделение металлопротеиназ, способствующих ремоделингу внеклеточного матрикса. Клетка перемещается в определенном направлении, «цепляясь» стресс-фибриллами поочередно за свои фокальные контакты.
Процесс клеточного движения регулируется малыми ГТФазами семейства Rho, такими как Cdc42, Rac, Rho. При этом в поляризованной движущейся мезенхимально клетке Cdc42 и Rac локализованы преимущественно на переднем крае и участвуют в образовании филоподий и ламеллиподий, а также в формировании первичных фокальных контактов. ГТФаза Rho в основном участвует в регуляции образования стресс-фибрилл и формировании натяжения в клетке. Все три ГТФазы взаимно регулируют активность друг друга, и баланс между ними очень важен для возможности клетки мигрировать и определять способ миграции.
Итак, основные признаки мезенхимального способа миграции клеток — это полимеризация актиновой сети, наличие фокальных контактов и деградация внеклеточного матрикса.
Коллективная миграция клеток
Увидеть коллективную миграцию клеток в культуре можно, нанеся ей экспериментальную «рану» (scratch assay). Тогда клетки будут стараться заполнить образовавшуюся пустоту, так же, как это происходит при ранении. Отдельные клетки в перемещающейся группе имеют мезенхимальные характеристики и движутся с помощью похожих механизмов, но между клетками в этой группе также образуются плотные кадгериновые контакты. Нередко при миграции такого ансамбля клеток появляются ведущие (или лидирующие) клетки, которые как бы тянут другие за собой.
Коллективная миграция клеток встречается не только при заживлении ран или во время эмбриогенеза, но также у агрессивных эпителиальных опухолей при инвазии. Сами по себе эпителиальные клетки не склонны к миграции. Однако они могут приобрести подвижный фенотип в результате эпителиально-мезенхимального перехода (ЭМП). После полноценного ЭМП клетки утрачивают эпителиальные маркеры (E-кадгерин, цитокератины), теряют апикально-базальную поляризацию и стабильные межклеточные контакты, но приобретают мезенхимальные маркеры (N-кадгерин, виментин, фибронектин). При коллективной миграции ЭМП происходит не полностью; межклеточные контакты сохраняются, но меняется их строение (например, из тангенциальных контакты становятся точечными или радиальными).
Амебоидное движение клеток
Для амебоидного типа движения характерно образование блебов. Формирование их регулируется ГТФазой Rho. При данном способе передвижения ведущую роль играет сокращение клетки, а не полимеризация актина. Клетка не формирует контактов с субстратом и перемещается очень быстро. Блебы обладают менее плотной внутренней структурой, чем псевдоподии, что в том числе и облегчает амебоидное передвижение.
При амебоидном движении тоже возникает поляризация клетки, только задний край отвечает за сокращение клетки и как бы подталкивает ее вперед. На переднем крае актиновый кортекс немного ослаблен, а из-за повышенного давления внутри клетки (за счет внешнего сжатия и/или возрастания сократимости актинового цитоскелета) она выбрасывает вперед блебы.
Это довольно быстрое передвижение, так как клетке не нужно прикрепляться к субстрату, она как бы перекатывается вперед. Почему же не все клетки передвигаются таким образом? Дело в том, что в этом случае клетка не выделяет металлопротеиназы, которые могли бы расчищать перед ней путь в матриксе. «Протиснуться» сквозь поры матрикса она не всегда может, в этом ее ограничивают размеры ядра. К тому же иногда клетке просто необходимо взаимодействовать с субстратом.
Миграционная пластичность опухолевых клеток
Для некоторых опухолевых клеток характерна пластичность — способность менять фенотип в ответ на внешние или внутренние сигналы без приобретения генетических мутаций. Особенно важна пластичность при метастазировании. Так, при выходе из первичного очага клетка должна преодолевать очень разные субстраты: пройти через пласт соединительной ткани, попасть к кровеносному сосуду, пройти через его мембрану, пережить кровоток, снова выйти из сосуда и пройти через матрикс, а затем оказаться в другой части организма. С похожими трудностями сталкиваются и нормальные клетки, например, иммунные.
Только мезенхимальное или только амебоидное движение не позволит преодолеть все эти препятствия. Поэтому клетки, обладающие способностью к пластичности, приобретают преимущества: они могут прибегать к полному или частичному ЭМП, а также к мезенхимально-амебоидному переходу.
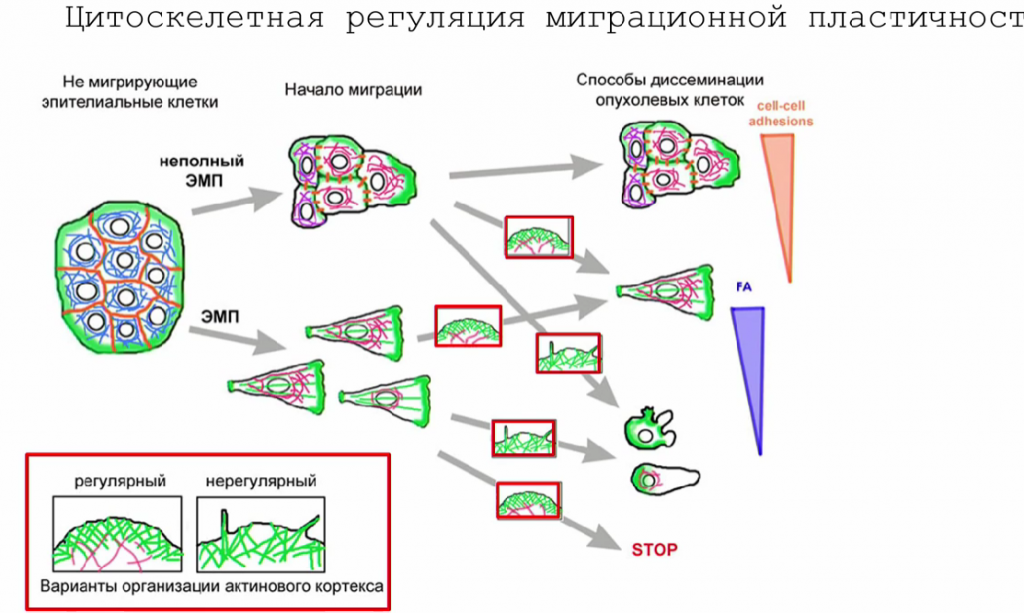
Опухолевые клетки обычно более пластичны, чем здоровые. Это видно даже по структуре их цитоскелета: такая клетка отличается большей готовностью к его перестройке для передвижения. Некоторые опухолевые клетки более пластичны, чем другие (особенно стволовые опухолевые клетки), и они могут стать «лидерами», которые ведут за собой других, или теми, кто формирует очаги вторичного роста. Такую пластичность Мария Ломакина и ее коллеги не раз демонстрировали в исследованиях, химически блокируя полимеризацию актина и другие процессы или заменяя субстрат на менее привычный для клеток. В результате клетки могут, например, переходить на амебоидный тип передвижения вместо мезенхимального.
«Одна и та же клетка за наши две с половиной минуты наблюдения попробовала разные способы. Она и распластывалась на субстрате, и формировала блебы, — комментирует Мария Ломакина. — Вторая решила, что ей по соседним клеткам удобнее бегать, когда она формирует блебы, а по стеклу удобнее, когда она мезенхимально передвигается».
Миграцию клеток в опухоли может стимулировать ее прогрессия (когда образуются новые мутантные клетки-клоны), воспаление и даже лечение — хирургическое вмешательство, радиотерапия, антиангиогенная терапия, применение ингибиторов митоза. Поэтому очень важно изучать особенности миграции клеток опухоли и искать «точки», на которые можно подействовать, чтобы предотвратить такое грозное осложнение опухолевого процесса, как формирование метастазов; или определять индивидуальный профиль лечения больных с онкологическими заболеваниями.
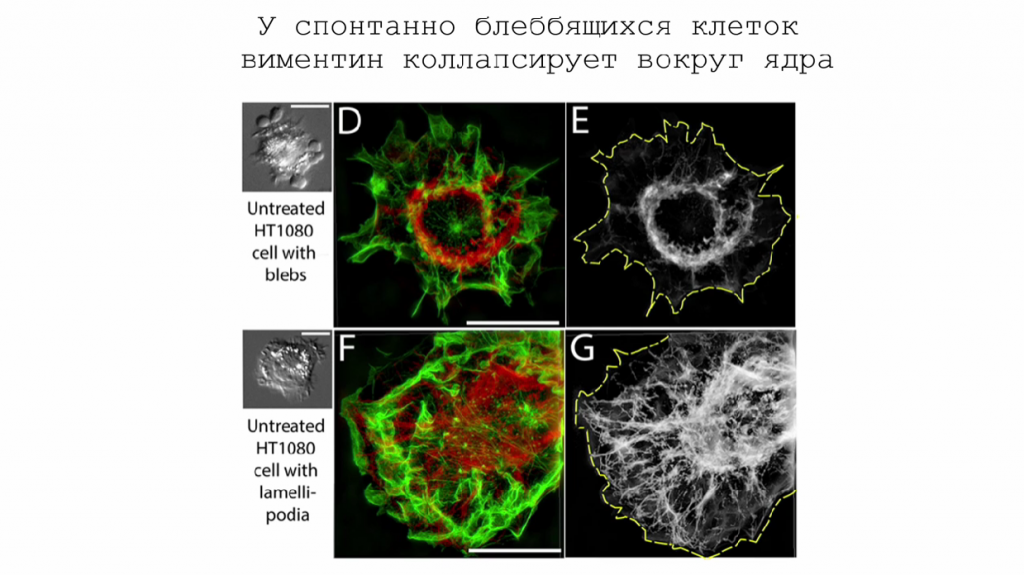
Так, в лаборатории механизмов канцерогенеза НМИЦ онкологии Н.Н. Блохина, в которой работает Мария Ломакина, были показаны изменения в морфологии виментиновых промежуточных филаментов на примере клеток фибросаркомы человека. Когда эти клетки двигаются амебоидным способом, виментин образует кольцевые структуры вокруг ядра, но при мезенхимальном движении промежуточные филаменты равномерно распределены по всему периметру клетки.
 Cells.2024 Jan 4;13(1):105. DOI:
10.3390/cells13010105
Cells.2024 Jan 4;13(1):105. DOI:
10.3390/cells13010105
Важность промежуточных филаментов в миграции клеток до сих пор была недооценена, но в последние годы ей уделяется особое внимание. В работе лаборатории механизмов канцерогенеза изучают роль виментиновых промежуточных филаментов в миграции.
При нокауте гена, кодирующего виментин, или внесении мутации, которая мешает сборке таких филаментов, клетки внешне нормальны. Однако они образуют меньше фокальных контактов. При помещении клеток в коллагеновый гель, отличающийся сложной 3D-структурой, контрольные фибробласты и фибробласты с нокаутом по виментину не перемещались в нем, а клетки с мутантным виментином активно мигрировали.
Сейчас в лаборатории проводят похожие эксперименты с опухолевыми клетками, и первичные данные показали, что клетки немелкоклеточного рака легкого в похожем эксперименте вели себя аналогичным образом, только клетки с мутацией в гене виментина полноценно переходили на амебоидный тип движения. Это значит, что виментин, возможно, участвует в мезенхимально-амебоидном переходе и стоит изучать его роль в процессе миграции и в дальнейшем, возможно, рассматривать в качестве новой «мишени» в рамках борьбы со злокачественным ростом, подвела итог Мария Ломакина.
Другие лекции блока «Клетка»
Гелина Копеина: «Без клеточной гибели терапия рака не сработает»
Виктор Татарский: «Все, что я вам рассказал, — это немножко ложь или сильное упрощение»

Вам будет интересно

 96
96
 0
0



 284
0
284
0